

Community hub
Recent from talks
Contribute something
Nothing was collected or created yet.
Xanthosoma
View on Wikipedia
| Xanthosoma | |
|---|---|

| |
| Xanthosoma roseum 'Elephant ear' | |
| Scientific classification | |
| Kingdom: | Plantae |
| Clade: | Tracheophytes |
| Clade: | Angiosperms |
| Clade: | Monocots |
| Order: | Alismatales |
| Family: | Araceae |
| Subfamily: | Aroideae |
| Tribe: | Caladieae |
| Genus: | Xanthosoma Schott[1] |

| |
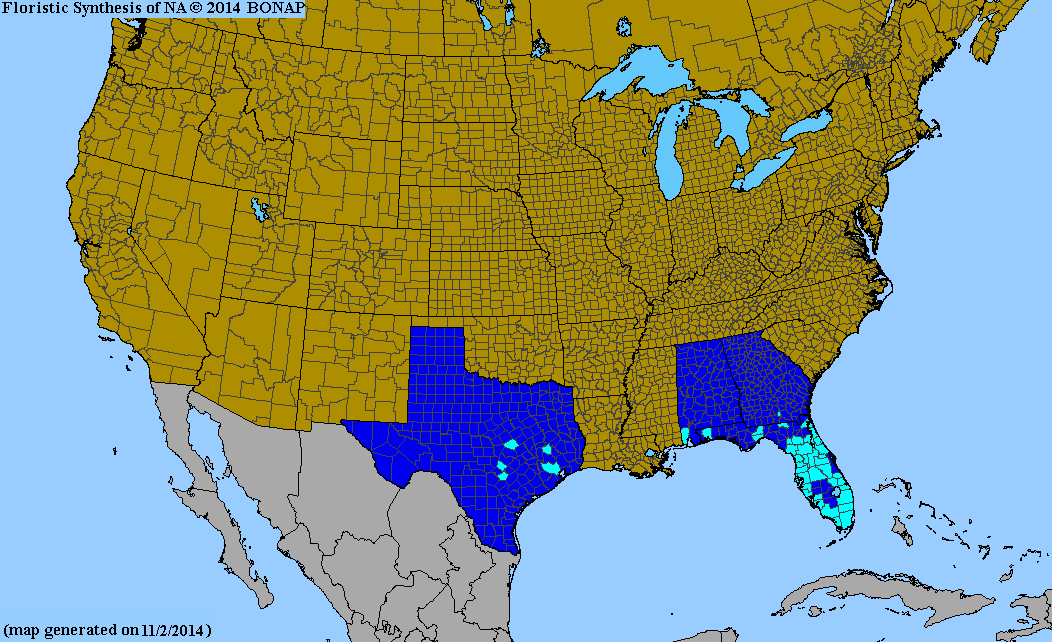
| Range of the genus Xanthosoma | |
| Synonyms[2] | |
| |
Xanthosoma is a genus of flowering plants in the arum family, Araceae. The genus is native to tropical America but widely cultivated and naturalized in other tropical regions.[2] Several are grown for their starchy corms, an important food staple of tropical regions, known variously as malanga, otoy, otoe, cocoyam (or new cocoyam), tannia, tannier, yautía, macabo, ocumo, macal, taioba, dasheen, quequisque, ʻape and (in Papua New Guinea) as Singapore taro (taro kongkong). Many other species, including especially Xanthosoma roseum, are used as ornamental plants; in popular horticultural literature these species may be known as ‘ape due to resemblance to the true Polynesian ʻape, Alocasia macrorrhizos, or as elephant ear from visual resemblance of the leaf to an elephant's ear. Sometimes the latter name is also applied to members in the closely related genera Caladium, Colocasia (taro), and Alocasia.
The leaves of most Xanthosoma species are 40–200 centimetres (16–79 inches) long, sagittate (arrowhead-shaped) or subdivided into three or as many as 18 segments. Unlike the leaves of Colocasia, those of Xanthosoma are usually not peltate- the upper v-notch extends into the point of attachment of the leaf petiole to the blade.
Reproduction
[edit]The inflorescence in Xanthosoma is composed of a spadix with pistillate flowers at the base, a belt of sterile flowers offered as a reward for pollinators in the middle and staminate flowers on the upper part. Prior to opening, the inflorescence is enclosed within a leaf-like spathe. When the inflorescence is ready to open, the upper part of the spathe opens and exposes the staminate area of the spadix; the basal area of the spathe remains closed, forming a spacious chamber (i.e., the spathe tube) that encloses the pistillate and sterile flowers (Garcia-Robledo et al. (2004, 2005a, 2005b)).
The inflorescences last for two nights and are protogynous in some, but not all species.[3] They change from the pistillate phase that attracts pollinators on the night it opens, to a staminate phase on the second night, when pollen is shed. When the inflorescence opens, it produces heat and releases a sweet scent attracting its pollinators, dynastine beetles (Cyclocephala spp.). Dynastines arrive covered with pollen from another inflorescence and remain in the spathe tube for 24 hours, pollinating the pistillate flowers as they feed on the sterile area of the spadix. On the second night, they come out of the tube and walk over the staminate flowers, getting covered with pollen, and then flying to a recently opened inflorescence nearby. (Garcia-Robledo et al. (2004, 2005a, 2005b)). Fruit maturation takes several months. Fruits start to develop within the shelter of the spathe tube. When the infructescence is mature, in some species, it arches back and downwards. In other species, it stays erect. Then, the tissue of the spathe tube rolls outwards, exhibiting the bright orange fruits and the velvety pink inner spathe surface.[4][5][6][3]
Taxonomy
[edit]Species
[edit]The following species are accepted:[2]
- Xanthosoma acutum E.G.Gonç. - French Guiana, Amapá State of Brazil
- Xanthosoma akkermansii (G.S.Bunting) Croat - Amazonas + Barinas States of Venezuela
- Xanthosoma aristeguietae (G.S.Bunting) Madison - Venezuela, northwestern Brazil
- Xanthosoma auriculatum Regel - northwestern Brazil
- Xanthosoma baguense Croat - northern Peru
- Xanthosoma bayo G.S.Bunting - Venezuela
- Xanthosoma belophyllum (Willd.) Kunth - Colombia, Venezuela, the Guianas; naturalized in Dominican Republic
- Xanthosoma bilineatum Rusby - Colombia
- Xanthosoma bolivaranum G.S.Bunting - Venezuela
- Xanthosoma brasiliense (Desf.) Engl. – Tahitian spinach - Lesser Antilles, Puerto Rico, Hispaniola, Trinidad & Tobago; naturalized in southern Brazil
- Xanthosoma brevispathaceum Engl. - Peru
- Xanthosoma caladioides Grayum - Panama
- Xanthosoma caracu K.Koch & C.D.Bouché – yautia horqueta - Puerto Rico, Dominican Republic
- Xanthosoma caulotuberculatum G.S.Bunting - Venezuela
- Xanthosoma conspurcatum Schott - Venezuela, Suriname, French Guiana
- Xanthosoma contractum G.S.Bunting - Bolívar State of Venezuela
- Xanthosoma cordatum N.E.Br. - Guyana, French Guiana
- Xanthosoma cordifolium N.E.Br. - Guyana
- Xanthosoma cubense (Schott) Schott - Cuba
- Xanthosoma daguense Engl. - Colombia, Ecuador
- Xanthosoma dealbatum Grayum - Costa Rica
- Xanthosoma eggersii Engl. - Ecuador
- Xanthosoma exiguum G.S.Bunting - Amazonas State of Venezuela
- Xanthosoma flavomaculatum Engl. - Colombia
- Xanthosoma fractum Madison - Peru
- Xanthosoma granvillei Croat & Thomps. - French Guiana
- Xanthosoma guttatum Croat & D.C.Bay - Valle del Cauca in Colombia
- Xanthosoma hebetatum Croat & D.C.Bay - Valle del Cauca in Colombia
- Xanthosoma helleborifolium (Jacq.) Schott – belembe silvestre - from Costa Rica south to central Brazil; naturalized in West Indies
- Xanthosoma herrerae Croat & P.Huang - Colombia
- Xanthosoma hylaeae Engl. & K.Krause - Colombia, Ecuador, Peru, Bolivia, northwestern Brazil
- Xanthosoma latestigmatum Bogner & E.G.Gonç. - Venezuela
- Xanthosoma longilobum G.S.Bunting - Venezuela
- Xanthosoma lucens E.G.Gonç - Rondônia
- Xanthosoma mafaffoides G.S.Bunting - Amazonas State of Venezuela
- Xanthosoma mariae Bogner & E.G.Gonç. - Peru
- Xanthosoma maroae G.S.Bunting - Amazonas State of Venezuela
- Xanthosoma maximiliani Schott - eastern Brazil
- Xanthosoma mendozae Matuda - México State in central México
- Xanthosoma mexicanum Liebm. - Chiapas, Oaxaca, Central America, Colombia, Venezuela
- Xanthosoma narinoense Bogner & L.P.Hannon - Colombia
- Xanthosoma nitidum G.S.Bunting - Venezuela
- †Xanthosoma obtusilobum Engl. - Mexico, probably extinct
- Xanthosoma orinocense G.S.Bunting - Amazonas State of Venezuela
- Xanthosoma paradoxum (Bogner & Mayo) Bogner - Colombia
- Xanthosoma pariense G.S.Bunting - Venezuela
- Xanthosoma peltatum G.S.Bunting - Venezuela
- Xanthosoma pentaphyllum (Schott) Engl. - Brazil
- Xanthosoma platylobum (Schott) Engl. - Brazil
- Xanthosoma plowmanii Bogner - Brazil
- Xanthosoma poeppigii Schott - Peru, Bolivia, northwestern Argentina
- Xanthosoma pottii E.G.Gonç. - Mato Grosso do Sul
- Xanthosoma puberulum Croat - Bolivia
- Xanthosoma pubescens Poepp. - Ecuador, Peru, Bolivia, northwestern Brazil
- Xanthosoma pulchrum E.G.Gonç. - Mato Grosso
- Xanthosoma riedelianum (Schott) Schott - southeastern Brazil
- Xanthosoma riparium E.G.Gonç. - Goiás
- Xanthosoma robustum Schott – capote - Mexico, Central America; naturalized in Hawaii
- Xanthosoma sagittifolium (L.) Schott (Syn. Xanthosoma atrovirens K.Koch & C.D.Bouché, Xanthosoma violaceum Schott)- arrowleaf elephant ear, tiquizque, macal, nampi, malanga or American taro[7] - Costa Rica, Panama, Venezuela, Colombia, Ecuador, Peru, Bolivia, Brazil; naturalized in West Indies, Africa, Bangladesh, Borneo, Malaysia, Christmas Island, Norfolk Island, some Pacific Islands, Alabama, Florida, Texas, Georgia,[8] Oaxaca[9]
- Xanthosoma saguasense G.S.Bunting - Venezuela
- Xanthosoma seideliae Croat - Bolivia
- Xanthosoma stenospathum Madison - Peru
- Xanthosoma striatipes (K.Koch & C.D.Bouché) Madison - Brazil, the Guianas, Venezuela, Colombia, Bolivia, Paraguay
- Xanthosoma striolatum Mart. ex Schott - French Guiana, northern Brazil
- Xanthosoma syngoniifolium Rusby - Bolivia, Argentina, Brazil
- Xanthosoma taioba E.G.Gonç. - Paraíba
- Xanthosoma tarapotense Engl. - Peru
- Xanthosoma trichophyllum K.Krause - Peru, Ecuador
- Xanthosoma trilobum G.S.Bunting - Amazonas State of Venezuela
- Xanthosoma ulei Engl. - northwestern Brazil
- Xanthosoma undipes (K.Koch) K.Koch – tall elephant's ear - widespread from Bolivia north to southern Mexico and West Indies
- Xanthosoma viviparum Madison - Peru, Ecuador
- Xanthosoma weeksii Madison - Ecuador
- Xanthosoma wendlandii (Schott) Schott (syn. Xanthosoma hoffmannii Schott, Xanthosoma pedatum Hemsl.) Oaxaca, Central America, Venezuela
- Xanthosoma yucatanense Engl. - Yucatán, Quintana Roo
Deprecated
[edit]- Phyllotaenium lindenii André (as X. lindenii (André) Engl.)
Etymology
[edit]The name is derived from the Greek words ξανθός (xanthos), meaning 'yellow', and σῶμα (soma), meaning 'body'. It refers to the stigma or yellow inner tissues.[10]
Uses
[edit]| Rank | Country | 2012 | 2013 | 2014 |
|---|---|---|---|---|
| 1 | 153782 | 185922 | 269590 | |
| 2 | 75132 | 84516 | 85607 | |
| 3 | 43000 | 43000 | 41110 | |
| 4 | 29200 | 30000 | 30960 | |
| 5 | 11692 | 23742 | 30000 | |
| 6 | 32595 | 29104 | 28180 | |
| — | World | 378952 | 423415 | 508079 |

Domestication of Xanthosoma species (especially X. sagittifolium but also X. atrovirens, X. violaceum, X. maffaffa and others) is thought to have originated in northern lowland South America, then spread to the Antilles and Mesoamerica. Today, Xanthosoma is still grown in all those regions, but is especially popular in Cuba, the Dominican Republic and Puerto Rico, where it is used in alcapurrias or boiled. It is grown in Trinidad and Tobago, Guyana and Jamaica to make the popular callaloo dish,[12] and in St. Kitts and Nevis to make tannia fritters.[13]
It is also grown in West Africa, now a major producer, where it can be used as a replacement for yams in a popular regional dish called fufu.
Xanthosoma is also grown as a crop in the Philippines.
Traditionally, Xanthosoma has been a subsistence crop with excess sold at local markets, but in the United States, large numbers of Latin American immigrants have created a market for commercial production. In general, production has yet to meet demand in some areas. In Polynesia, Alocasia macrorrhizos (‘ape) was considered a famine food, used only in the event of failure of the much preferred taro (kalo) crop.[14] After having been introduced to Hawaii in the 1920s from South America, Xanthosoma has naturalized and has become more common than A. macrorrhizos, and has been given the same name, ʻape.
The typical Xanthosoma plant has a growing cycle of 9 to 11 months, during which time it produces a large stem called a corm, this surrounded by smaller edible cormels about the size of potatoes. These cormels (like the corm) are rich in starch. Their taste has been described as earthy and nutty, and they are a common ingredient in soups and stews. They may also be eaten grilled, fried, or puréed. The young, unfurled leaves of some varieties can be eaten as boiled leafy vegetables or used in soups and stews, such as the Caribbean callaloo.
Flour made from Xanthosoma species is hypoallergenic.[15]
Gallery
[edit]-
 Inflorescence of Xanthosoma sagittifolium
Inflorescence of Xanthosoma sagittifolium -
 Inflorescence of X. daguense
Inflorescence of X. daguense -
 Beetle pollination in X. daguense
Beetle pollination in X. daguense -
 Infructescence of X. poeppigii (Peruvian Amazonas); X. daguense (Western Cordillera of Los Andes, Colombia)
Infructescence of X. poeppigii (Peruvian Amazonas); X. daguense (Western Cordillera of Los Andes, Colombia)




References
[edit]- ^ "Genus: Xanthosoma Schott". Germplasm Resources Information Network. United States Department of Agriculture. 2003-07-09. Retrieved 2012-07-13.
- ^ a b c Kew World Checklist of Selected Plant Families
- ^ a b Valerio, C. E. (1988), "Notes on the phenology and pollination of Xanthosoma wendlandii (Araceae) in Costa Rica" (PDF), Rev. Biol. Trop., 36: 55–61
- ^ Garcia-Robledo, Carlos; et al. (2004), "Beetle pollination and fruit predation in Xanthosoma daguense (Araceae)", Journal of Tropical Ecology, 20 (4): 459–469, doi:10.1017/S0266467404001610, S2CID 85768260
- ^ Garcia-Robledo, Carlos; et al. (2005a), "Equal and opposite effects of floral offer and spatial distribution on fruit production and pre-dispersal seed predation in Xanthosoma daguense (Araceae)", Biotropica, 37 (3): 373–380, Bibcode:2005Biotr..37..373G, doi:10.1111/j.1744-7429.2005.00049.x, S2CID 86329238
- ^ Garcia-Robledo, Carlos; et al. (2005b), "Geographic Variation and Succession of Arthropod Communities in Inflorescences and Infructescences of Xanthosoma (Araceae)", Biotropica, 37 (4): 650–656, Bibcode:2005Biotr..37..650G, doi:10.1111/j.1744-7429.2005.00082.x, S2CID 45182954
- ^ Lim, T. K. (2015). "Xanthosoma sagittifolium". Edible Medicinal and non Medicinal Plants. pp. 498–509. doi:10.1007/978-94-017-9511-1_15. ISBN 978-94-017-9510-4.
- ^ Biota of North America Program, 2013 county distribution map
- ^ García-Mendoza, A. J. & J. Meave del Castillo. 2011. Divers. Florist. Oaxaca 1–351. Universidad Nacional Autónoma de México, Ciudad Universitaria
- ^ Quattrocchi, Umberto (2000). CRC World Dictionary of Plant Names. Vol. IV R-Z. Taylor & Francis US. p. 2849. ISBN 978-0-8493-2678-3.
- ^ "Production of Yautia (Cocoyam) by countries". UN Food & Agriculture Organization (FAO). 2014. Archived from the original on July 13, 2011. Retrieved 22 April 2016.
- ^ Caribbean Islands. Lonely Planet. 2021. p. 243. ISBN 978-1-78701-673-6.
- ^ Caribbean Islands. Lonely Planet. 2021. p. 682. ISBN 978-1-78701-673-6.
- ^ Abbott, Isabella Aiona. (1992). Lā'au Hawai'i : traditional Hawaiian uses of plants. [Honolulu, Hawaii]: Bishop Museum Press. p. 5. ISBN 0-930897-62-5. OCLC 26509190.
- ^ Vaneker, K. The Pomtajer. Page 216 In: Friedland, S. R., Ed. Vegetables: Proceedings of the Oxford Symposium on Food and Cooking 2008: Volume 26 of Proceedings of the Oxford Symposium on Food and Cookery. Oxford Symposium, 2009.
{kind=link}
External links
[edit] Media related to Xanthosoma at Wikimedia Commons
Media related to Xanthosoma at Wikimedia Commons Data related to Xanthosoma at Wikispecies
Data related to Xanthosoma at Wikispecies- Xanthosoma spp. at Purdue University, Center for New Crops & Plants Products
- Distribution and names of edible aroids Archived 2005-12-20 at the Wayback Machine
Xanthosoma
View on GrokipediaDescription
Vegetative Morphology
Xanthosoma species are herbaceous, perennial plants that grow from underground corms or tubers, typically reaching heights of 1 to 3 meters. They exhibit a robust, erect habit, often acaulescent in younger stages but developing a thick, fleshy stem up to 1 meter tall in mature individuals. The stems are subterranean and rhizomatous in structure, with cylindrical internodes up to 7 cm in diameter, moderately covered by brown fibers, and capable of profuse branching in older plants. These plants produce a milky sap containing calcium oxalate crystals, which can cause skin and mucous membrane irritation upon contact.[1][6][7] The leaves are a defining feature, emerging in a rosette from the crown of the central corm and characterized by their large size, ranging from 40 to 200 cm in length. Leaf shapes vary across the genus, including sagittate, cordate, peltate, and pedatisect forms. Leaf blades are typically sagittate or arrow-shaped, with a sharp apex and deep basal lobes, measuring 40–113 cm long and 30–70 cm broad or more, depending on species and conditions; they are typically non-peltate, though some species have peltate leaves, distinguishing most Xanthosoma from related genera like Colocasia which usually have peltate leaves. Petioles are long, often exceeding 2 meters, ribbed, and thick, supporting the erect or slightly inclined blades that display parallel venation with simple veins converging toward the apex. Blade texture is sub-coriaceous, and colors are predominantly green, though some species exhibit variegation or pigmentation.[8][9][3][10] Underground structures consist of a central starchy corm, which serves as the primary propagation organ and is enclosed by dry, scale-like leaves forming an outer husk. The corm has a lumpy exterior with rough ridges and brown skin, surrounding an inner edible portion of white, pink, or yellow flesh rich in digestible starch; however, the main corm is often acrid and less preferred for consumption compared to secondary structures. Swollen offshoots known as cormels develop from lateral buds on the corm or rhizomes, varying in size, shape, and number per plant, while fibrous roots extend from the base to anchor and absorb nutrients. In cultivated species, these corms and cormels are valued for their edibility after proper processing to mitigate oxalate content.[3][11][9] Morphological variations occur across species, reflecting adaptations and cultivar diversity. For instance, Xanthosoma sagittifolium features prominently sagittate leaves often described as heart-shaped due to their basal lobes, with petioles and veins showing color variations such as green or purplish hues among accessions. In contrast, Xanthosoma violaceum displays purple-tinged foliage and bluish leaf undertones, enhancing its ornamental appeal while maintaining the typical sagittate form. These differences in leaf shape, coloration, and corm pigmentation (e.g., yellow-fleshed cultivars) highlight the genus's variability, though all share the non-peltate leaf attachment and erect growth.[12][13][3]Inflorescence Structure
The inflorescences of Xanthosoma emerge from the leaf axils, typically numbering 1–3 (up to 6 in some species) per plant, and consist of a central spadix enclosed by a single spathe. The peduncle supporting the inflorescence measures 5–60 cm in length, varying by species and environmental conditions.[10][14] The spadix, which is shorter than the spathe and measures 3.7–30 cm long, is divided into three distinct zones: a basal pistillate zone with female flowers, a middle sterile zone consisting of staminodes, and an apical staminate zone with male flowers. This zonation supports a protogynous flowering sequence in many species, where the pistillate zone becomes receptive prior to anthesis of the staminate zone, promoting cross-pollination. The sterile zone features thickened staminodia that produce food bodies, serving as a reward for dynastine beetle pollinators, while the overall spadix exhibits thermogenesis and fragrance emission to attract these insects.[15][16][17] The spathe is boat-shaped, comprising a persistent lower tube (ovoid to oblong, 4–10 cm long) and a deciduous upper blade (10–20 cm long overall for the spathe), with the tube often green, purple, or reddish on the exterior and the blade typically white or creamy inside. During anthesis, the spathe blade opens over two nights, with color shifts such as from green to pale yellow in the tube or purple blotching in species like X. cerrosapense, facilitating pollinator access before wilting.[10][14] Flowers are unisexual and naked, lacking perianth; pistillate flowers form syncarps that develop into cylindroid, many-seeded berries, while staminate flowers consist of 4–6 connate stamens forming a truncate-obpyramidal synandrium without free filaments, distinct from some related Araceae genera. The stigma is notably yellow and discoid to hemispheric-discoid, often 3–4-lobed and producing a jelly-like exudate when receptive, a trait reflected in the genus name derived from Greek for "yellow body."[14][15][18]Taxonomy
Etymology
The genus name Xanthosoma is derived from the Ancient Greek words xanthos (ξανθός), meaning "yellow," and sōma (σῶμα), meaning "body," referring to the yellow inner tissues found in some species of the genus.[19] The name was coined by the Austrian botanist Heinrich Wilhelm Schott in his publication Meletemata Botanica, issued in 1832, as part of his systematic contributions to the classification of the Araceae family during the early 19th-century exploration of tropical flora.[20] Schott's work built on contemporary botanical efforts to organize Neotropical aroids based on reproductive and vegetative traits. Common names for Xanthosoma species vary widely across regions, reflecting their cultural significance in tropical agriculture and cuisine; prominent examples include malanga and yautia in the Caribbean, cocoyam and tannia in parts of Africa and Asia, and elephant ear in English-speaking areas for their large foliage. In Brazil, the term taioba is commonly used, particularly for the edible leaves of certain species.[21] These vernacular names often trace their origins to indigenous languages of tropical America, such as Taino (for yautia) and Arawak (for malanga), indicating pre-Columbian uses by native peoples in Central and South America.[22]Classification History
The genus Xanthosoma was established by Heinrich Wilhelm Schott in 1832, based on material from tropical America, with the type species X. sagittifolium (L.) Schott.[20] Initially, the genus encompassed several synonyms, including Acontias Schott (1832) and Cyrtospadix K.Koch (1855), which were later reduced as distinct genera were refined within the Araceae family. In the early 20th century, Adolf Engler classified Xanthosoma within the subfamily Colocasioideae, tribe Colocasieae (subtribe Caladiinae) in his comprehensive treatment of Araceae in Das Pflanzenreich (1905–1920), emphasizing its tuberous habit and inflorescence structure as key diagnostic features shared with allies like Caladium.[23] This placement highlighted Xanthosoma as part of a core group of Neotropical aroids, though Engler's system predated molecular insights and relied heavily on morphology. Subsequent refinements in the mid-20th century maintained this tribal affiliation, evolving into the modern tribe Caladieae.[24] Major taxonomic revisions in the 1980s and 2000s, led by Thomas B. Croat and collaborators, significantly expanded understanding of Xanthosoma's diversity, recognizing approximately 50–75 species based on field collections and morphological analyses across the Neotropics.[3] These efforts addressed longstanding gaps in South American taxa, with Croat's work emphasizing sectional divisions and ecological adaptations. A pivotal 2017 revision of Central American species by Croat, Delannay, and Ortiz treated 18 taxa and described seven new species, such as X. cerrosapense and X. hammelii, refining distributions and resolving synonyms from earlier works.[10] As of 2024, additional species such as X. alpayacuense have been described from Colombia, further increasing the recognized diversity of the genus.[25] Species delimitation in Xanthosoma remains debated due to frequent hybridization and the proliferation of cultivated forms, many of which have been misidentified under X. sagittifolium, complicating wild-type distinctions.[1] Recent efforts include the 2023 neotypification of X. blandum Schott by Croat and De Dijn, using historical illustrations from Schott's Icones to stabilize its application amid ambiguous type material from Brazil.[6] Molecular phylogenetic studies confirm Xanthosoma's position within the tribe Caladieae of subfamily Aroideae, closely related to Caladium through shared plastid genome features and anastomosing laticifers, while aligning more distantly with Alocasia in the broader Aroideae clade based on multi-locus analyses.[26][27]Accepted Species
The genus Xanthosoma currently comprises approximately 200 accepted species, all native to tropical regions of the Americas from Mexico southward to northern Argentina and Bolivia.[20] These species are primarily herbaceous perennials in the family Araceae, with the type species designated as Xanthosoma sagittifolium (L.) Schott, a widely cultivated plant known for its large sagittate leaves and edible cormels used in traditional agriculture across the Neotropics.[1] Recent taxonomic revisions have significantly expanded the recognized diversity through detailed field studies and herbarium examinations, emphasizing distinctions in vegetative and reproductive morphology.[28] Among the accepted species, several stand out for their economic or ecological significance. Xanthosoma sagittifolium serves as the primary cultivated taxon, valued for its starchy cormels that form a dietary staple in many tropical communities.[1] Xanthosoma violaceum Schott is notable for its edible corm and violet-tinged inflorescences, distributed from Central America to northern South America.[10] Ornamental forms with rose-colored petioles, often associated with the name Xanthosoma mafaffa Schott (a synonym of X. sagittifolium), are found in wet forests of Central and South America.[29] [10] Geographic variation is evident, with species such as Xanthosoma daguense Engl. restricted to montane forests in northern South America, particularly Colombia, and Xanthosoma hannoniae Croat & L.P. Hannon endemic to premontane habitats in Central America.[30] Several names previously recognized as distinct species have been deprecated as synonyms based on overlapping morphological traits and cultivation-induced variation. Common examples include:| Deprecated Name | Synonym Of | Reason for Deprecation |
|---|---|---|
| Xanthosoma atrovirens K. Koch & C.D. Bouché | X. sagittifolium | Represents a dark-green variant of the cultivated type species, lacking consistent diagnostic differences.[1] |
| Xanthosoma caracu K. Koch & C.D. Bouché | X. sagittifolium | Cultivated form with minor petiole color variation, merged due to artificial selection artifacts.[1] |
| Xanthosoma nigrum (Vell.) Stellfeld | X. sagittifolium | Misidentification based on dark leaf markings, resolved through inflorescence comparison.[31] |
| Xanthosoma roseum Schott | X. undipes (K. Koch & C.D. Bouché) K. Koch | Applied to ornamental selections with pink petioles, but identical in key floral traits.[10] [32] |
| Xanthosoma croatianum Croat | X. mexicanum Engl. | Based on incomplete specimens; morphological overlap confirmed in revision.[10] |
| Xanthosoma pilosum K. Krause | X. mexicanum Engl. | Synonymized due to variable pubescence not warranting separation.[10] |
| Xanthosoma hoffmannii Engl. | X. wendlandii Schott | Historical misapplication from herbarium errors in leaf dissection.[10] |
| Xanthosoma jacquinii Schott | X. undipes (K. Koch & C.D. Bouché) K. Koch | Illegitimate name; traits align with X. undipes peduncle morphology.[10] |
| Caladium helleborifolium (Jacq.) W.Wight | X. helleborifolium (Jacq.) Schott | Pre-generic transfer; no differences in spadix structure.[10] |
| Xanthosoma mendozae Matuda | X. wendlandii Schott | Regional variant with indistinguishable inflorescence features. |