Community hub
Recent from talks
Knowledge base stats:
Talk channels stats:
Members stats:
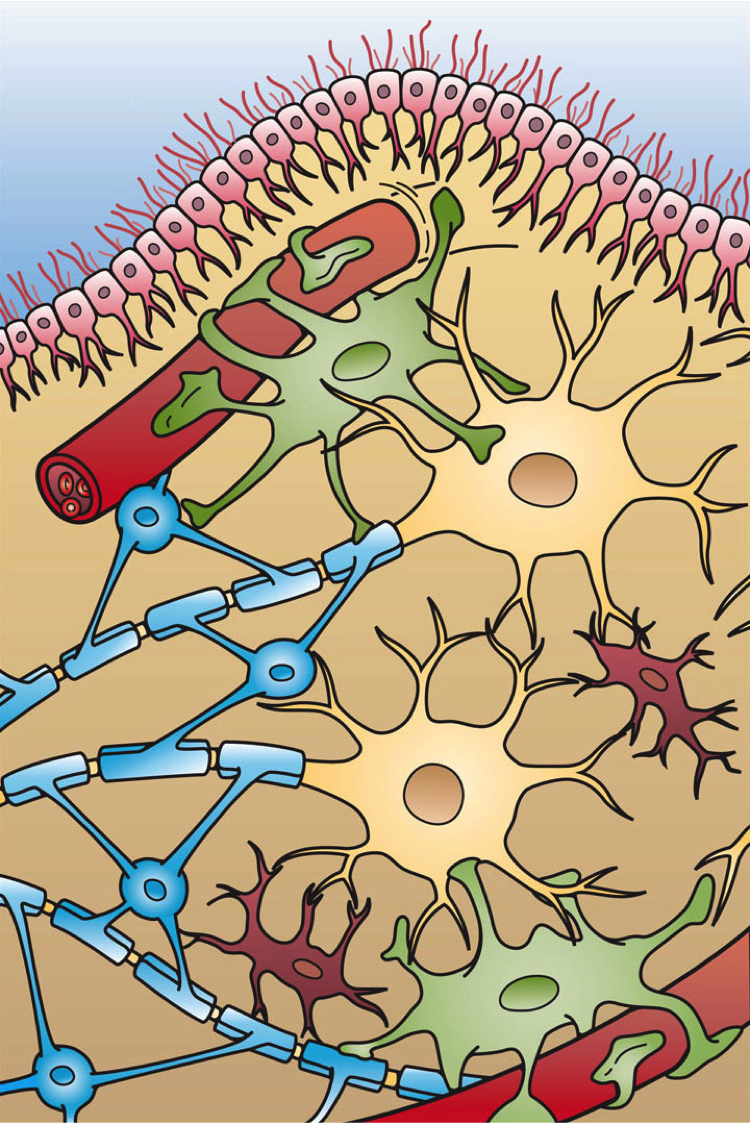
Glia
Glia, also called glial cells (gliocytes) or neuroglia, are non-neuronal cells in the central nervous system (the brain and the spinal cord) and in the peripheral nervous system that do not produce electrical impulses. The neuroglia make up more than one half the volume of neural tissue in the human body. They maintain homeostasis, form myelin, and provide support and protection for neurons. In the central nervous system, glial cells include oligodendrocytes (that produce myelin), astrocytes, ependymal cells and microglia, and in the peripheral nervous system they include Schwann cells (that produce myelin), and satellite cells.
Glia have four main functions:
They also play a role in neurotransmission and synaptic connections, and in physiological processes such as breathing. While glia were thought to outnumber neurons by a ratio of 10:1, studies using newer methods and reappraisal of historical quantitative evidence suggests an overall ratio of less than 1:1, with substantial variation between different brain tissues.
Glial cells have far more cellular diversity and functions than neurons, and can respond to and manipulate neurotransmission in many ways. Additionally, they can affect both the preservation and consolidation of memories.
Glia were discovered in 1856, by the pathologist Rudolf Virchow in his search for a "connective tissue" in the brain. The term derives from Greek γλία and γλοία "glue" (English: /ˈɡliːə/ or /ˈɡlaɪə/), and suggests the original impression that they were the glue of the nervous system.
Derived from ectodermal tissue.
Microglia are specialized macrophages capable of phagocytosis that protect neurons of the central nervous system. They are derived from the earliest wave of mononuclear cells that originate in the blood islands of the yolk sac early in development, and colonize the brain shortly after the neural precursors begin to differentiate.
These cells are found in all regions of the brain and spinal cord. Microglial cells are small relative to macroglial cells, with changing shapes and oblong nuclei. They are mobile within the brain and multiply when the brain is damaged. In the healthy central nervous system, microglia processes constantly sample all aspects of their environment (neurons, macroglia and blood vessels). In a healthy brain, microglia direct the immune response to brain damage and play an important role in the inflammation that accompanies the damage. Many diseases and disorders are associated with deficient microglia, such as Alzheimer's disease, Parkinson's disease and ALS.
Hub AI
Glia AI simulator
(@Glia_simulator)
Glia
Glia, also called glial cells (gliocytes) or neuroglia, are non-neuronal cells in the central nervous system (the brain and the spinal cord) and in the peripheral nervous system that do not produce electrical impulses. The neuroglia make up more than one half the volume of neural tissue in the human body. They maintain homeostasis, form myelin, and provide support and protection for neurons. In the central nervous system, glial cells include oligodendrocytes (that produce myelin), astrocytes, ependymal cells and microglia, and in the peripheral nervous system they include Schwann cells (that produce myelin), and satellite cells.
Glia have four main functions:
They also play a role in neurotransmission and synaptic connections, and in physiological processes such as breathing. While glia were thought to outnumber neurons by a ratio of 10:1, studies using newer methods and reappraisal of historical quantitative evidence suggests an overall ratio of less than 1:1, with substantial variation between different brain tissues.
Glial cells have far more cellular diversity and functions than neurons, and can respond to and manipulate neurotransmission in many ways. Additionally, they can affect both the preservation and consolidation of memories.
Glia were discovered in 1856, by the pathologist Rudolf Virchow in his search for a "connective tissue" in the brain. The term derives from Greek γλία and γλοία "glue" (English: /ˈɡliːə/ or /ˈɡlaɪə/), and suggests the original impression that they were the glue of the nervous system.
Derived from ectodermal tissue.
Microglia are specialized macrophages capable of phagocytosis that protect neurons of the central nervous system. They are derived from the earliest wave of mononuclear cells that originate in the blood islands of the yolk sac early in development, and colonize the brain shortly after the neural precursors begin to differentiate.
These cells are found in all regions of the brain and spinal cord. Microglial cells are small relative to macroglial cells, with changing shapes and oblong nuclei. They are mobile within the brain and multiply when the brain is damaged. In the healthy central nervous system, microglia processes constantly sample all aspects of their environment (neurons, macroglia and blood vessels). In a healthy brain, microglia direct the immune response to brain damage and play an important role in the inflammation that accompanies the damage. Many diseases and disorders are associated with deficient microglia, such as Alzheimer's disease, Parkinson's disease and ALS.