Respect all members: no insults, harassment, or hate speech.
Be tolerant of different viewpoints, cultures, and beliefs. If you do not agree with others, just create separate note, article or collection.
Clearly distinguish between personal opinion and fact.
Verify facts before posting, especially when writing about history, science, or statistics.
Promotional content must be published on the “Related Services and Products” page—no more than one paragraph per service. You can also create subpages under the “Related Services and Products” page and publish longer promotional text there.
Do not post materials that infringe on copyright without permission.
Always credit sources when sharing information, quotes, or media.
Be respectful of the work of others when making changes.
Discuss major edits instead of removing others' contributions without reason.
If you notice rule-breaking, notify community about it in talks.
Do not share personal data of others without their consent.
Separation, such as a spore detaching from a sporogenous cell. From Latin abscissio, breaking off.[4]
abstriction
A method of spore formation in fungi characterized by abjunction and then abscission. Spores are produced in a sporogenous filament. In abjunction, they are then separated by transverse walls or septa, with the result spores are grouped in short chains. In abscission, successive portions are cut off and released.[5]
acidophilous
acidophilic
Organisms that can grow in high-acidity environments; in mycology, lichens that can grow in peaty soil or on acidic tree bark.[6]
acropetal
A chain of conidia where new spores are formed at the apex, with the oldest at the base. Compare with basipetal.[7]
acropleurogenous
The acropleurogenous Trichothecium roseum, with conidia on sides and at apex
pleuroacrogenous
Formed at the apex and along the sides; e.g. conidia of Trichothecium.[8]
adiaspore
Conidium of Emmonsia parva. A large spore that increases notably in size, but does not divide. Upon being inhalted in the lungs of humans and animals, can cause adiaspiromycosis disease. From Gr. a-, without, dia, separating.[9]
adnate
attached, adherent
Adhering; attached to the stipe throughout its width, esp. of lamellae or tubes. Compare with free.[10]
aero-aquatic fungi
Aquatic fungi that grow in water, but spread their spores via air.[11]
An agaric, with gills on the underside of the pileus
A member of the order Agaricales; a mushroom or toadstool. Contains many iconic and highly studied fungi. Agarics have a macroscopic (human-visible) fleshy basidioma with a clearly differentiated stipe (stalk), pileus (cap), and lamellae (gills) on the underside of the pileus.[13]
Fusion between branches of hyphae to make a network.[17]
annellidic
Magnified view of Scopulariopsis brevicaulis, its annellides, and the ring-shaped annellations emanating from them
A type of blastic conidiogenesis. The conidiogenous cell (also called an annellide) produces a basipetal sequence of conidia called annelloconidia or annellospores. The distal end is marked by ring-shaped bands called annellations. Examples include many aquatic Hyphomycetes, Venturia inaequalis, and Microascus brevicaulis.[18]
annular
1. Ring-like; an arrangement in the style of a ring.
Having no stem or pedicel, such as a pileus lacking a stipe and directly attached to the substrate. Found in sporangia of Perichanea for example. From Gr. podos, foot.[25]
apothecium
pl. apothecia, discocarp
A type of fruiting body that, at maturity, opens to expose the hymenium of asci; commonly assumes shape of a cup or saucer. The stipe is sometimes lichenized. From Gr. apotheke, storehouse.[26]
Flattened, or closely applied; e.g. of scales or fibers.[28]
appressorium
pl. appresoria
A swelling at the end of a germ tube or other hypha that adheres to the surface of its host and penetrates it with infection hyphae. Characteristic of plant pathogenic fungi such as Phytophthora and Colletotrichum, as well as parasites of animals such as Entomophthora.[29]
aquatic fungi
hydrofungi
Fungi that live in water. Sometimes subdivided into freshwater-living fungi and marine fungi for saltwater-living fungi. Freshwater fungi include: Many zoosporic fungi of Chytridiomycota, Chytridiales, and Saprolegniales; some saxicolous lichens of Lichinaceae that live on submerged rocks; aquatic hyphomycetes or Ingoldian fungi, a group of mitosporic fungi that live in freshwater with branched or sigmoidial spores, often growing on dead leaves in streams; smuts of aquatic plants; and certain yeasts.[30]
archicarp
The cell, hypha, or coil of Ascomycetes that later becomes the ascoma, or part of it.[31]
ardella
A small spot-like apothecium, as in the lichen Arthonia. From Latin ardere, to sprinkle.[32]
A delimited space on a surface, separated from others by fissures or cracks.[33]
arthric
thallic-arthric
A method of thallic conidiogenesis that converts a hyphal element into a conindium (also called an arthrospore), fragmenting the hypha. Common in many basidiomycetes. Compare with holothallic. From Gr. arthron, joint.[34]
asc-
asco-, ascidi-
A prefix meaning sac, bladder, or ascus. From Gr. askós, vessel, bag, or wineskin.[35]
A phylum of fungi characterized by the presence of an ascus, a sac-like structure where ascospores are produced. The largest group of fungi. Includes cup fungi or Discomycetes; most dermatophytes; the mycobiont part of most lichens; powdery mildews; and fungi that produce truffles.[37]
Spores formed in the developing ascus, generally as a result of karyogamy (nuclear fusion) followed by meiosis. Commonly, four haploid daughter nuclei divide to make eight haploid nuclei, around which eight ascospores are created by depositing wall material around them, a process sometimes called free cell formation.[38]
The reproductive cell of ascomycetes; where ascospores form and are contained. While sometimes traditionally restricted to only sexual reproduction, purely asexual mitosporic fungi have since been classified as having asci as well (List of mitosporic Ascomycota).[39]
assimilative
vegetative, somatic
Hyphae related to growth, nutrition, and asexual reproduction as opposed to sexual reproduction; the soma.[40]
A spore close in form to a zygospore, but developed asexually (parthogenesis). Found in many species of Mucorales and some species of Entomophthorales, such as Entomophthora muscae. The structure that contains it is called an azygosporangium.[42]
A phylum of fungi. Generally defined by sexual reproduction via basidiospores formed from a basidium, although a few purely anamorphic basidiomycetes exist. Typically mycelial, although some are yeasts and some are dimorphic. Basidiomycetes include earth balls; earthstars; false truffles; jelly fungi; many mushrooms; polypores; puffballs; most rusts; most smuts; and stinkhorns.[45]
A chain of conidia in which new spores are formed at the base and the oldest are at the apex. Compare with acropetal.[48]
blastic
One of the two basic forms of conidiogenesis, with thallic conidiogenesis. Characterized by the enlargement of the conidia initial before it is delimited by a septum. In holoblastic, both inner and outer walls of the blastic conidiogenous cell contribute to the formation of the conidium; in enteroblastic, only the inner walls enlarge and contribute. Monoblastic is from a single conidiogenous locus; polyblastic is when a conindiogenous cell has multiple conidiogenous loci.[49]
An iconic variety of mushroom characterized by fleshy fruit bodies and a poroidhymenophore (marked by pores rather than the gill-marked cap of agarics). They are often edible. Boletes are members of the order Boletales.[51]
A type of asexual cellular multiplication. A small outgrowth or bud from a parent cell enlarges and eventually separates from the parent cell. Typical in yeasts and many molds. Sometimes divided by how many buds are made, with monopolar, bipolar, and multipolar budding for one/two/many respectively.[52]
bulbil
A compact multicellular propagule, with its thin-walled, undifferentiated cells produced in acropetal succession from the hyphae. Superficially resembles plant tissue at maturity (pseudoparenchymatous) such as raspberries, hence the resemblance to bulbils in botany. Found in certain basidiomycetes; characteristic of aero-aquatic fungi such as Bulbillomyces farinosus, where they grow on leaves or tree branches previously submerged in water.[53]
Combining forms meaning "fruit". From Gr. karpós, fruit.[55]
capsule
Magnification of the yeast Cryptococcus neoformans; the capsules are the circular outer borders of the yeast cells.
A clear, gelatinous covering or sheath around the cell wall of certain yeasts, e.g. Cryptococcus, and some ascospores such as those of Sordaria fimicola.[56]
catenate
Catenulate
Arranged in chains or end-to-end series. Catenulate is the diminutive form for small chains. From Latin catena, a chain.[57]
A 200x magnification of Candida albicans, including a chlamydospore
An asexual thick-walled spore developed from hyphae, generally for perennation rather than dissemination. From Gr. chlamys, cloak, -ydos, spore.[61]
A phylum of fungi. Informally known as chytrids. Characterized by chitinous cell walls and reproduction via zoospores, which are usually uniflagellate in the posterior although rarely polyflagellate. Many are saprobes that live in freshwater (see aquatic fungi) or the soil.[62]
cirrus
cirrhus; spore horn
A curl-like tuft; usually refers to a tendril-like mass of forced-out spores.[63]
Clamp connection formation between two nuclei (one in green, the other orange)
clamp, fibula
A hyphal outgrowth that makes a connection during cell division between the resulting two cells by fusion. Generally associated with mycelium of Basidiomycetes.[64]
clavate
Narrowing at the base and then thickening; club-shaped.[65]
An artificial taxon for mitosporic fungi with pycnidial and acervular states, i.e. they form their spores in an internal cavity. From Gtk. koilos, hollow.[67]
Hyphae that lack septa and are multi-nucleate; seen in Zygomycota and Oomycota. From Gr. kytos, cell. Distinct from a synctium, a multinucleate structure resulting from fusion of protoplasts.[69]
Producing conidia. Generally used as "conidiogenous cell", fertile cells that produce conidia; or "conidiogenous locus", for the particular point on a hypha or a cell where conidia are generated.[74]
conidioma
pl. conidiomata
Any multi-hyphal, conidia-bearing structure. An umbrella term that includes various traditional conidia-bearing structures such as a synnema, a sporodochium, an acervulus, or a pycnidium.[75]
A cobweb-like cortina of Cortinarius claricolor
Of agarics, the remnants of the partial veil, frequently web-like, that covers mature gills. Thin and evanescent; eventually breaks up and disappears.[81]
Crust-like or having a hard surface layer, e.g. in a sporocarp; in lichens, a thallus firmly fixed to the substratum by the whole of their lower surface, generally lacking a distinct lower cortex.[84]
cyphella
pl. cyphellae
A break in the lower cortex of a lichen thallus which appears as a cup-like structure or marking. Characteristic of Sticta. From Gr. kyphella, the hollow of an ear.[85]
A parasitic fungus that focuses on tissue with keratin (skin, hair, nails) of humans and animals. Associated with the genera Epidermophyton, Microsporum, and Trichophyton.[90]
The skin condition caused by a dermatophyte infection.[91]
dichotomous
Forking into roughly equal pairs, often repeatedly, e.g. in hyphal branching. Compare with monopodial.[92]
dictyospore
(1) An amerospore with no intersecting septa; (2) a didyomospore with a single intersecting septa; (3) two sample phragmospore patterns with multiple septa subdividing the spore; (4) a dictyospore with a brick wall-like muriform pattern of intersecting septa
Divided by intersecting septa in more than one plane, both transverse and longitudinal; a muriform spore. Found in Alternaria alternata as an example.[93]
didymospore
A two-celled spore divided by a single septum. Usually applied to mitosporic fungi. Compare with amerospore and phragmospore.[94]
A class of saucer-shaped and cup-shaped Ascomycetes in which the hymenium is exposed at maturity; one in which the fruit body is an apothecium (discocarp). Prominent members include Pezizales, Helotiales, Ostropales, and Lecanorales.[99]
disjunctor
An evanescent connective, consisting of either a cell or cell wall material. It develops through the pores of septal lamellae of adjoining conidia in a chain, before later fragmenting and releasing the conidia.[100]
In hyphae of basidiomycete fungi, parenthesomes (1) "cap" a dolipore septum; (2) the cell wall; (3) swells around the septal pore to form a barrel-shaped ring. Perforations in the parenthesome allow cytoplasm to flow between (4) and (5).
A septum found in Basidiomycetes that flares out in the middle to form a channel.[101]
Having sharply pointed spines, e.g. of spores. Its diminutive is echinulate, for delicate spines. Sometimes synonymous with spinose. From Gr. echinos, hedgehog.[102]
effete
Past the bearing period; overmature, e.g. a fruiting body that has emptied its spores.[103]
effuse
Expanded; stretched out flat, e.g. a film-like growth. From Latin fundere, to pour.[104]
An organism that lives within a plant; in mycology, specifically fungi that live within plants but do not show external signs or damage to the plants. This is usually endomycorrhizial fungi in root systems and asymptomatic fungi in aerial plant parts[108]
Having a short existence; soon disappearing, e.g. of a veil, an annulus, etc. From Latin evanscere, to disappear.[117]
eucarpic
eucarpous
Thalli where reproductive structures only develop on limited portions during fructication. The rest of the thallus remains in its original assimilative function. Compare holocarpic. If they have one sporangium, they are monocentric; if they have several sporgangia, they are polycentric. The vast majority of fungi are eucarpic.[118]
A fairy ring
Mushrooms or puffballs forming in a circle, started from a central mycelium in the soil. Fairy rings expand with time if undisturbed, and are generally associated with basidiomycetes.[122]
falcate
Falciform
Curved, like a sickle or scythe. From Latin falx and falcis, sickle.[123]
A chemical change caused by enzymes of living organisms. In mycology, typically yeasts and molds performing anaerobic breakdowns such as changing carbohydrates to carbon dioxide and alcohol. Part of the creation of fermented foods and fermented drinks (most notably alcoholic beverages).[124]
filament
A fine thread. Commonly used for hyphal threads or other simple thread-like fungal structures. From Latin filum, thread.[125]
filamentous
filamentose
1. Thread-like, or composed of filaments.
2. In lichens, when the photobiont forms in threads surrounded by hyphae of the mycobiont; the fungi forms the outer structure. (e.g. Coenogonium, Cystocoleous, Racodium)[126]
Fluffy or cottony; byssoid. Common among many colonies of fungi. Its diminutive form is flocculose, for a small and delicate floccose element. From Latin floccus, a tuft of wool.[131]
2. Of lichens, foliose lichens, characterised by flattened leaf-like thalli.[135]
foot cell
basal cell
1. A hyphal cell that supports a sporogenous cell or thallus, specifically the support of the conidiophore in Aspergillus and the macroconidium of Fusarium.
Panelluses stipticus, an example of luminiscient fungi (long exposure photograph)
A phosphorescent light emitted by moist, decaying wood; one of the most famous forms of fungal bioluminescence.[137]
A substance able to kill fungi, especially at low concentration. From Latin caedere, to kill. Generally used for chemical substances rather than heat, light, radiation, etc. which are called sterilization instead.[142]
A kingdom of organisms in Eukaryota. Fungi are distinguished based on their morphology, nutritional modes, and ecology. Typical traits are that they lack plastids, are osmotrophic (absorb nutrients from their environment), are never phagotrophic, lack an ameboid pseudopodal phase, cell walls are composed of a chitin-glucan complex, mitochondria have flattened cristae and perioxomes are nearly always present, and are mostly non-flagellate; flagella when present lack mastigonemes. From Gr. sphongos, σπόγγος, sponge.[147]
1. Another term for chlamydospores, sometimes restricted to specifically spores that broke free from the mycelium to be dispersed, often via water. From Latin gemma, "jewel"
Germ tubes forming in Candida albicans, at 600x magnification
A short hypha that sprouts from the germ pore during germination. If conditions are favorable, will form a full mycelium. In plant pathogenic fungi, forms an appressorium, from which the infective hypha spread and penetrate the tissues of the host.[159]
Downy mildew infiltrating cells of the plant Arabidopsis thaliana, at 400x magnification. The stained dark blue spheres are the haustoria.
A hyphal branch for absorption of food. It penetrates the wall of the living host cell. Found in certain parasitic fungal plant pathogens, such as Phytophthora.[170]
3. A condition where a flagellum possess two rows of tripartite tubular hairs.[172]
heteromerous
In lichens, when the thallus is stratified. The photobiont is distributed in a compact layer, and the hyphal tissue of the fungi are separated into an outer rind and an inner stratum. From Gr. hetero, different, and meros, part.[173]
A scar after a spore is discharged; was previously the point of attachment of a basidiospore to the sterigma. From Latin hilum, mask.[175]
hoary
Covered thickly with silk-like hairs, especially of a pileus or stipe, often grayish or white.[176]
holocarpic
Having all of the thallus converted into the fruiting body at the end of maturation; compare eucarpic. The somatic and reproductive phases do not coexist as a result. Examples include Olpidiaceae and Synchytriaceae.[177]
Term describing the whole fungus throughout all its morphs (states, phases). Generally includes a single teleomorph phase and one or more anamorph phases.[178]
holothallic
A method of thallic conidiogenous in which a hyphal element is converted as a whole into a single conidium. All the cell wall layers are involved. Compare with arthric. From Gr. holos, whole.[179]
homoiomerous
In lichens, when the photobiont is are distributed uniformly or at random throughout the thallus. From Gr. homoios, similar.[180]
The spore-bearing fruiting surface of Ascomycetes and Basidiomycetes. An aggregation of asci or basidia in a layer (palisade) mixed with other sterile cells.[186]
An artificial class of mitosporic fungi that reproduces asexually. Traditionally subdivided into Agonomycetales, Hyphomycetales, Stilbellales, and Tuberculariales.[190]
hypnospore
resting spore
A thick-walled resting spore; germinates only after a lapse of weeks or months. From Gr. hypnos, sleep.[191]
hypo-
Prefix meaning "under" or "lower". From Gr. hypo.[192]
1. (Of lichens) The first growth of the hyphae, before differentiation has taken place. Usually of a crustaceous lichen's earliest stage, lacking a cortex and photobiont. Often persists as a colored layer on the bottom of the later stages of maturation.
2. (of slime molds) a thin layer on the surface not used up in sporangial development.[193]
Not operculate; lacking an operculum. Generally refers to asci or sporangia that discharge spores by an irregular apical opening or pore instead. Found in various Discomycetes and most other ascomycetes.[194]
A propagule for lichen growth; can both increase the surface area of a lichen and become detached from the thallus to disperse the lichen. Isidia are structures larger than soredia, and contain both the mycobiont and the photobiont. They are usually cylindrical, but can also be verrucose, clavate, and other shapes.[196]
A gill; a hymenium-covered vertical plate, generally of an agaric. Attaches to the stipe in a variety of ways, including: free, unattached; adnate, attached; adnexed, a partial attachment; sinuate, similar to adnate but with an S-shaped curve; decurrent, attached and also running further down the surface of the stipe. From Latin lamina, a thin plate.[203]
Lactarius quietus exuding cream-colored latex from gills upon being cut
A milk-like juice that flows from some fungi when cut or damaged, as in Lactarius.[207]
Crustose lichens can grow on surfaces hostile to most other lifeforms, such as concrete walls and bare boulders.
A dual organism that arises from an association of fungi (mycobiont) and algae or cyanobacteria (photobiont). The two coexist in a mutualistic relationship as partners; the resulting lifeform differs markedly from its isolated components.[211]
Rhizocarpon geographicum (map lichen) in Switzerland
A technique for studying the exposure age of rock surfaces based on the size and diameter of lichen thalli. Used extensively in glaciology. Most frequently uses map lichen for dating. Lichenometry has been used to date Easter Island moai among other elements.[216]
An illustration of ectomycorrhiza
A layer of hyphae covering the roots of ectomycorrhizal plants, generally trees. Connected to a Hartig net on the inside, and extramatrical hyphae on the outside.[227]
A series of two successive nuclear divisions that reduces the number of chromosomes by one-half, going from a diploid state to a haploid one. The last part of the sexual phase in many fungi, following plasmogamy and karyogamy. Contrast with mitosis.[231]
merosporangium
pl. merosporangia
A sporangium that is a cylindrical outgrowth from the swollen tip of a sporangiophore. A chain-like row of sporangiospores are produced there. Characteristic of Mucorales; also found in some other zygomycetes.[232]
A fungus of very small size, usually with microscopic sporocarps.[241]
monokaryotic
monocaryotic
Cells having a single nucleus each; having genetically identical haploid nuclei (monokaryon or haplont). Found, for example, in the mycelium of Agaricales.[242]
monopodial
A type of branching where there is a persistent main axis from which branches split from, one at a time, often in alternate or spiral series. Often uses acropetal succession. Compare with dichotomous.[243]
A division of fungi broken off from Zygomycota in the 2010s that includes many of the molds, microscopic fungi. Includes Mucorales, the largest and most closely studied order of zygomycetes.[248]
Amanita muscaria (fly agaric), an iconic mushroom
Large agarics, or other fleshy fungi such as boletes. Commonly divided into mushrooms (human-edible) and toadstools (inedible).[251]
Hemitrichia serpula, commonly called pretzel slime, a true slime mold
Myxomycetes
True slime molds. No longer categorized as fungi, but were formerly categorized as such in older literature, and still studied in mycology; now considered part of Amorphea. Since reclassified as Myxogastria. From Gr. myxa, slime.[267]
Riesling grapes afflicted by Botrytis cinerea (gray rot), causing a color change
A condition from the mold Botrytis growing on overripe grapes. Used in the production of certain sweet wines.[269]
Water molds. Traditionally considered fungi, but now classed as closer to algae. Part of the Chromista kingdom. Contain cellulose in their cell walls rather than chitin, and coenocytic hypae rather than septate hyphae.[274]
400x magnification showing oospores of Hyaloperonospora parasitica, a downy mildew
The resting spore formed as a result of fertilization of the oosphere. Has a thick, resistant wall. Subdivided into centric, subcentric, subeccentric, and eccentric oospores by its structure.[276]
A small rounded elevation. Generally refers to an elevation on the wall of the sporangium which on breaking serves as the exit point of zoospores and planogametes.[282]
paraphysis
pl. paraphyses
A sterile upward-growing, basally-attached hypha in a hymenium, especially in ascomata of ascomycetes where they are generally filiform and clavate. The free ends frequently converge toward the ostiole and make a structure called an epithecium over the asci. Part of the hamathecium. Has a number of minor variants distinguished by names such as paraphysoid, pseudoparaphysis, pseudophysis, dikaryoparaphysis, and hyphidium.[283]
A sample agaric-type basidioma in (A) the early development stage, and (B) after the body is fully expanded. (1) is the universal veil, the outer layer protecting the developing basidioma; (2) is the partial veil, which covers the gills; (3) are cap scales, remnants of the universal veil; (4) is the volva, another remnant of the universal veil, but at the base of the basidioma; (5) is the annulus, a ring-like mark on the stipe that is a remnant of the partial veil, and whose overhanging tissue may become a cortina.
A layer of tissue that joins the stipe to the edge of the pileus in agarics, covering the gills during hymenium development. May become an annulus or a cortina after development is complete.[284]
pectinate
Resembling the teeth of a comb, e.g. of the margin of a pileus.[285]
1. Conidiogenous cell growth where a conidiophore or germ tube grows through a preexisting pore.
2. Of a columella, extending throughout the entire length, from the base of the gleba through to the peridium's apex. Found in gastromycetes such as Podaxis. Also called a stipe-columella or dendritic columella.[289]
Feeding by engulfing food; extending a pseudopod or other protoplasmic extension, as seen in protozoa. Never seen in true fungi (Eumycota), but some Pseudofungi use this in addition to osmocytosis, e.g. Myxomycetes.[292]
A type of conidiogenous cell, bottle-shaped, that produces blastic conidia (phialospores) in basipetal succession. The philade itself does not increase in length. The most common conidiogenous cell among conidial fungi. From Gr. phiale, jar.[293]
phialospore
An asexual spore formed from the tip of a phialide. Formed by abstriction.[294]
A tropic phenomenon driven by light, where growth curves toward or against light stimuli. For example, sporangiphores of Pilobus and Phycomyces direct themselves toward light.[296]
A group of forms alike in morphology. Often means a group of organisms that are potentially interbreeding. In plant pathology, it means a group with similar preferences in plants targeted; a race may be adapted to target only a single cultivar of a plant. Tracking the history of an organism is phylogeny. From Old Italian razza, generation.[298]
The fusion of two cells or plasmodial cytoplasms, resulting in the nuclei juxtaposed and a dikaryon formed. In many sexual phases of fungi, the first step which precedes karyogamy (nuclear fusion) and meiosis.[303]
plectenchyma
A thick, packed tissue of twisted hyphae formed during basidiocarp development as it enlarges and generative hyphae begin to differentiate. From Gr. plektos, to twist, and enchyma, infusion.[304]
Any structure that serves to spread the organism, both via propagation of new organisms as well as vegetatively increasing a single organism's size. In fungi, generally spores, bulbils, fragments of mycelium, isidia, soredia, and others.[309]
pseudo-
Prefix meaning "false", from Gr. pseudos.
pseudoidia
Separated hyphal cells capable of germination.[310]
A geniculate or zig-zag holoblastic extension of a conidiogenous cell from sympodial conidiogenous cell development. Such cells having a rachis are called rachiform. From Gr. ráchis, axis, spine.[320]
racket cell
A hyphal cell having a swelling at one end, resembling a tennis racket; found in dermatophytes.[321]
A slender, tapered structure of anucleate filaments bearing a superficial resemblance to a plant root, as it is extended by the thallus of chytrids as a feeding organ. Generally part of a root system-like aggregation of branched hyphae.[325]
A strand or cord of mycelium, often with a dark-colored rind surrounding a central core of colorless cells, penetrating a soft substratum or between portions of it (e.g. between bark and wood). Unlike a mycelial cord, features distinct tissue, unlike "normal" hyphae. Functions as organ of absorption of nutritive substances. Seen in some Agaricales and Gasteromycetes.[326]
A sister group of quasi-fungi that lack chitinous cell walls. Traditionally considered chytrids, but have since been separated and reclassified in the 2010s. Phagotrophs rather than osmotrophes. Rozella, a group of obligate endoparasites, is possibly the earliest diverging lineage of fungi.[328]
A hardened, often rounded, mass of hyphae, normally having no spores. May give rise to a fruit body, a stroma (as in ergot), conidiophores, or mycelium. Can be a store of nutrients as part of perennation; can reinitiate vegetative growth after conditions have improved and a season has passed.[336]
scutate
scutiform
Like a shield or round plate in shape. From Latin scutum, shield.[337]
A fruiting body with the appearance of an unopened agaric or bolete. The margin of the pileus does not break free of the columella-stipe, generally seen in vertical development (e.g. Podaxis pistillaris).[338]
1. Growing old. From Latin senescere, to grow old.
2. The degeneration over time that makes indefinite propagation of certain fungi cultures impossible, especially in isolation such as a lab environment.[341]
Edged with teeth, like a comb or saw, e.g. of gills; its diminutive form is serrulate.[344]
sexual phase
The phase of the fungal life cycle when sexually reproductive cells and organs are formed. This term is preferred over sex in animals, as the fungal process is not equivalent.[345]
Acrasis rosea, a cellular slime mold
A eukaryotic lifeform that spreads via spores. Has both ameba (cellular slime molds) and plasmoidial (multi-nucleate) varieties. No longer categorized as fungi, but were formerly categorized as such in older literature. Cellular slime molds are part of Acrasiomycota or Dictyosteliomycota; plasmodial slime molds were traditionally part of the class Myxomycota, since renamed Myxogastria. From Gr. myxa, slime.[346]
1. Loose smut of barley, caused by Ustilago nuda
A class of destructive plant diseases, generally of cereal grasses, caused by parasitic fungi of Ustilaginomycetes. Distinguished by transformation of plant organs permeated by hyphae into a dark mass of spores.
2. Smut-causing fungi themselves in Ustilaginomycetes. Characterized by being host-specific endophytes. Its spores are called ustilospores.
A propagule for vegetative lichen growth; a combination of phycobiont algae wrapped by mycobiont hyphae, and produced on a lichen thallus. Has the appearance of a powdery granule. From Latin soredium, a small heap.[350]
A heap of spores. Fruiting structure in certain fungi, including the spore mass of rusts, but also Acrasidae and Synchytriaceae. From Gr. soros, heap.[351]
spawn
Mycelium used for starting fungal cultures, especially mushrooms; e.g. bricks of manure interlaced with mycelia.[352]
A reproductive structure in fungi. Can result from both sexual and asexual processes.[360]
spore wall
The layered wall defining a spore. Considered to have five layers. From within to outwards: the thin interior endosporium, the thick episporium, the exosporium (or tunic), the perisporium, and ectosporium, although the outermost two layers are fleeting and can be absorbed back into the perisporium and exosporium.[361]
A mass of vegetative hyphae where fertile hyphae, fruiting bodies, and spores are produced. Common among ascomycetes and anamorphic fungi; a few Pucciniales have them as well.[370]
The sexual state (or perfect state) of a fungus whose spores are produced by meiosis, i.e. characterized by ascomata or basidiomata.[378]
thallic
One of the two basic forms of conidiogenesis, with blastic conidiogenesis. Characterized by the conidia initial being delimited by one or more septa before it begins enlargement. The result is that the conidium is differentiated from the whole cell. By comparison, in blastic conidiogenesis, enlargement occurs within the cell before being delimited later.[379]
1. A layer of hyphae in the central part of an agaric running from the underside of the cap to the lamella, often supporting the hymenium. Sometimes called a hymenophoral trama to distinguish it from the second definition.
2. In old literature, any fleshy part of the pileus, although this use has since been discouraged.[382]
Arranged in a single row or series. Generally used to differentiate how phialides are arranged in species of Aspergillus; in uniseriate, they are directly on the conidial head, contrasted with biseriate where phialides rest on intermediate outgrowths of sterile cells called metulae.[394]
The white patches on the caps of these Amanita muscaria mushrooms are cap scales, remnants of the universal veil.
A layer of tissue covering the basidioma during its early development, in agarics and Gasteromycetes. As the pileus grows, the veil is broken, with its upper remnants becoming cap scales, and the lower section becoming the volva.[395]
Warty; having rounded bumps. Verruculose is the diminutive version for delicate or small warts.
verticillate
Having parts in rings (verticils); whorled. Develops due to branching in which branches or pedicels are borne at the same level on the hypha, and grow obliquely upward with respect to the central axis. Named after the conidiophores of Verticillium, but appears in other fungi such as the sporangiophore of Actinomucor. From New Latin verticillatus, arranged in a verticil.[397]
Fungi that digest wood. Mostly basidiomycetes, although a few ascomycetes also possess this ability. Generally categorized into brown rot, which digests a tree's cellulose and hemicellulose but not its lignin; white rot, which can also digest lignin; and soft rots, which are similar to brown rots in attacking cellulose and hemicellulose, but require moist wood and available nitrogen, e.g. from nearby soil. Two other notable types are dry rot (a slight misnomer, as some dampness is still required), a brown rot caused by Serpula lacrimans; and wet rot, several other species including cellar fungus.[404]
Preferring a dry habitat, or at least capable of subsisting in one. Rare among fungal species, but fungi with this capability can be exceptionally common, such as Aspergillus and Penicillium whose spores can be found in nearly any soil sample.[405]
A glossary of mycology is a specialized reference compiling definitions of technical terms used in the scientific study of fungi, encompassing vocabulary related to their morphology, reproduction, taxonomy, ecology, and pathology.[1] These glossaries serve as essential tools for mycologists, students, and researchers to interpret scientific literature and describe fungal characteristics precisely.[2]Prominent examples include A Glossary of Mycology by Walter H. Snell and Esther A. Dick, first published in 1957 and revised in 1971 by Harvard University Press, which features over 6,000 entries with illustrations of key fungal structures like hyphae, spores, and fruiting bodies, and was acclaimed for its utility in clarifying complex terminology across scientific and medical fields.[3] Another authoritative work is Dictionary of the Fungi (10th edition, 2008) by Paul M. Kirk, Paul F. Cannon, David W. Minter, and J. A. Stalpers, published by CABI, containing more than 21,000 entries that provide comprehensive coverage of fungal taxa at genus, family, and order levels, along with biographical notes on notable mycologists and references to influential publications.[2] These resources highlight the evolution of mycological nomenclature, adapting to advances in fungal classification and molecular techniques.The importance of mycological glossaries lies in standardizing language amid the diversity of fungi—estimated at 2.2 to 3.8 million species, with approximately 155,000 (about 4–7%) formally described as of 2025[4][5]—and facilitating interdisciplinary applications in areas like agriculture, medicine, and environmental science.[2] Terms such as hypha (a filamentous fungal cell), mycelium (a network of hyphae), and basidiocarp (a spore-producing fruiting body) are foundational, enabling precise communication about fungal life cycles and interactions, including symbiotic relationships like mycorrhizae.[1] Ongoing updates to these glossaries reflect taxonomic revisions, such as the integration of phylogenetic data, ensuring relevance in contemporary mycology.[2]
Taxonomy and Classification
Major Phyla and Groups
The kingdom Fungi is a monophyletic group within the eukaryotic supergroup Opisthokonta, sister to the lineage including animals and amoeboid protists like Nucleariida and Filasterea, with an inferred ancestral zoosporic (flagellated) form.[6] Modern fungal classification recognizes eight major phyla based on molecular phylogenies, emphasizing traits such as hyphal septation, reproductive structures, and ecological roles, while excluding non-fungal groups like slime molds.[7] These phyla encompass diverse morphologies from unicellular yeasts to complex multicellular forms, with approximately 160,000 described species representing only a fraction of estimated fungal diversity.[5]Early-diverging phyla include Rozellomycota (also known as Cryptomycota), comprising intracellular parasites such as Rozella species that infect organisms like oomycetes, characterized by reduced morphology, zoosporic stages with digitiform protrusions, and chitin in some life phases but often lacking cell walls during the trophic stage.[6]Blastocladiomycota features zoosporic fungi with complex thalli, including saprotrophic and parasitic species in aquatic and moist terrestrial environments. Neocallimastigomycota consists of anaerobic, flagellated fungi inhabiting the rumens of herbivores, adapted for lignocellulose degradation without mitochondria. Chytridiomycota, another basal zoosporic lineage, features flagellated zoospores and simple thalli (unicellular or rhizomycelial), with three classes—Chytridiomycetes, Monoblepharidomycetes, and Neocallimastigomycetes—encompassing saprobes, parasites, and gut inhabitants of herbivores; this phylum diverged approximately 750–1,000 million years ago and includes amphibian pathogens like Batrachochytrium dendrobatidis.[7][8] Oomycota, historically misclassified as fungi due to filamentous growth and zoosporic reproduction, are actually stramenopiles (related to brown algae) with cellulose cell walls and oosporesexual reproduction, serving as plant pathogens but excluded from Fungi.[6]The paraphyletic Zygomycota, an outdated phylum defined by coenocytic (aseptate) hyphae and zygospore formation, has been split based on genomic and phylogenetic evidence into Mucoromycota and Zoopagomycota.[7]Mucoromycota includes three subphyla—Mucoromycotina (saprobic molds like Mucor), Mortierellomycotina (soil saprobes), and Glomeromycotina (arbuscular mycorrhizal symbionts with ~370 species forming ancient plant associations without observed sexual reproduction)—characterized by coenocytic hyphae, sporangia, and sometimes chitosan in cell walls.[6][9] Zoopagomycota encompasses subphyla like Zoopagomycotina (animal and amoebal parasites), Kickxellomycotina, and Entomophthoromycotina (insect pathogens), with coenocytic hyphae, zygospores, and a retained centrosome, often lacking dolipore septa.[7]The subkingdom Dikarya unites Ascomycota and Basidiomycota, defined by a prolonged dikaryotic phase (two unfused nuclei per cell) and septate hyphae, representing the majority of terrestrial fungi with complex life cycles.[6]Ascomycota, the largest phylum (~66,000 species, comprising about 40–45% of described fungi), produces sexual spores in sac-like asci (typically eight ascospores), with three subphyla—Taphrinomycotina (simple yeasts), Saccharomycotina (budding yeasts like Saccharomyces), and Pezizomycotina (molds and lichens)—featuring Woronin bodies in septa and diverse roles as saprobes, pathogens, and symbionts.[7][10]Basidiomycota (~36,000 species) generates spores on club-shaped basidia, divided into subphyla Pucciniomycotina (rusts), Ustilaginomycotina (smuts), and Agaricomycotina (mushrooms and puffballs), with dolipore septa (parenthesome-lined pores) in many members and ecological importance in wood decay, mycorrhizae, and plant diseases.[6][10]Historically, fungal taxonomy distinguished Eumycota (true fungi with chitinous walls and filamentous growth) from Myxomycota (slime molds with amoeboid plasmodia and cellulose walls, now classified separately in Amoebozoa).[11] Outdated terms like Gymnomycota (once encompassing naked slime molds) and Chitomycetes (early reference to chitin-walled fungi) persist as artifacts of pre-molecular classifications, superseded by phylogenomic approaches tracing major divergences to 550–400 million years ago following land plant colonization.[11]
Life Cycle Stages
In fungal biology, life cycle stages encompass the developmental phases from sporegermination to reproduction, often involving both asexual and sexual processes that contribute to taxonomic identification. These stages are particularly emphasized in the context of pleomorphic fungi, where morphological variation across phases complicates classification. The holomorph concept integrates all stages of a fungus's life cycle, providing a unified view for nomenclature and systematics.[12]The holomorph denotes the complete fungus, encompassing both asexual and sexual reproductive structures as well as vegetative phases. It represents the entirety of the organism's biological cycle, allowing mycologists to consider the full developmental spectrum rather than isolated forms. In contrast, the anamorph specifically refers to the asexual stage, characterized by mitotic spore production such as conidia, which facilitates rapid dispersal and colonization without genetic recombination.[12][13]The teleomorph describes the sexual stage, involving meiosis and karyogamy to produce genetically diverse spores like ascospores or basidiospores, which is crucial for evolutionary adaptation and species delineation. When a fungus exhibits multiple distinct asexual forms, additional anamorphs are termed synanamorphs, reflecting morphological diversity within the anamorphic phase, often observed in complex species like those in Ascomycota. This multiplicity can arise from environmental influences or genetic factors, aiding survival in varied niches.[14][15]Fungal reproduction integrates mitosis and meiosis, with mitosis dominating vegetative growth and asexual propagation, while meiosis ensures genetic variability in sexual cycles. A key event is karyogamy, the fusion of haploid nuclei from compatible mating types, which restores diploidy and precedes meiosis. In Ascomycota, karyogamy occurs within the ascus, followed by meiosis to yield four haploid nuclei that undergo an additional mitotic division, resulting in eight ascospores. Similarly, in Basidiomycota, karyogamy in the basidium leads to meiosis, producing four haploid basidiospores externally. These processes highlight the dikaryotic phase common in many fungi, where unfused nuclei coexist before karyogamy.[16][17]Many fungi exhibit dimorphic life cycles, alternating between unicellular yeast-like forms and multicellular hyphal structures in response to environmental cues such as temperature. For instance, pathogenic species like Histoplasma capsulatum grow as mycelia at ambient temperatures but switch to yeast forms at mammalian body temperature (37°C), facilitating infection. This dimorphism enhances adaptability, with the yeast phase often linked to parasitism and the hyphal phase to saprotrophic or environmental persistence.[18]Pleomorphic life cycles extend this variability, involving more than two morphological forms across the full cycle, including multiple anamorphs or transitions beyond simple dimorphism. Such pleomorphism is prevalent in groups like Erysiphales, where fungi produce both teleomorphic fruiting bodies and diverse anamorphic states, allowing exploitation of different ecological roles. This complexity historically challenged taxonomy, as forms were often described separately until molecular tools linked them.[19]Historically, terms like Discomycetes referred to a class of Ascomycota characterized by apothecial fruiting bodies, such as cup- or disc-shaped asci, encompassing operculate species now integrated into Pezizomycotina. Anamorphic groups were classified as Coelomycetes, denoting fungi producing conidia within enclosed pycnidia or acervuli, and Hyphomycetes, which form conidia openly on hyphae without specialized structures. These terms, from the era of Deuteromycota (fungi imperfecti), underscored asexual diversity before sexual-asexual linkages were established, influencing early mycology but largely superseded by phylogenetic classifications.[20][21]
Cellular and Structural Features
Hyphae, Septa, and Mycelium
Hyphae are the fundamental filamentous units that constitute the vegetative body of most fungi, typically measuring 2–10 μm in diameter and elongating through apical extension at the tip. These tubular structures branch repeatedly, enabling the fungus to explore and absorb nutrients from its substrate, and they form the basis of the fungal thallus in multicellular species. In contrast to unicellular yeasts, hyphal growth allows for polarized expansion driven by cytoskeletal elements and vesicle transport to the apex.[12][22]A mycelium represents the interconnected network of hyphae that collectively forms the main body of a fungus, often spreading extensively underground or within substrates to facilitate resource acquisition and colonization. This mat-like structure can span vast areas, such as in forest soils where it interconnects plantroots, and it exhibits dynamic behaviors like directed growth toward nutrient sources. Mycelia vary in density and pigmentation, from diffuse white mats to compact, pigmented aggregates, depending on environmental conditions and species.[23][24]Specialized mycelial aggregates include rhizomorphs and mycelial cords, which are linear, cord-like structures formed by the bundling and fusion of hyphae, often with a central core of vessel hyphae for efficient nutrienttransport. Rhizomorphs, typically found in wood-decay fungi like Armillaria species, enable long-distance translocation of water and nutrients through soil, resembling root-like organs with an outer sheath of tightly packed hyphae. These structures enhance fungal foraging and invasion capabilities, allowing growth over distances of several meters. Mycelial cords are similar but generally less organized, serving comparable roles in resource distribution within the mycelial network.[25][26]Septa are cross-walls that divide hyphae into compartments, with fungal hyphae classified as either septate or coenocytic based on their presence and regularity. Septate hyphae, common in Ascomycota and Basidiomycota, feature periodic septa that partition the filament into uninucleate or multinucleate cells while allowing cytoplasmic continuity through pores. Coenocytic (or aseptate) hyphae, prevalent in Mucoromycota and Zoopagomycota and some Chytridiomycota, lack septa or have them only sporadically, resulting in a continuous multinucleate protoplasm that supports rapid cytoplasmic streaming and growth. This organization influences fungal physiology, with coenocytic forms enabling faster expansion but potentially higher vulnerability to damage.[27][28][29]In Basidiomycota, septa often take the form of dolipore septa, characterized by a barrel-shaped swelling around a central pore that is typically plugged by a septal pore cap composed of membrane-bound vesicles. This structure maintains cellular integrity while permitting selective nucleocytoplasmic exchange, and it is associated with the dolipore septum's role in preventing organelle migration between compartments. Dolipore septa are a diagnostic feature of many higher basidiomycetes, contributing to their complex dikaryotic life cycles.[30][31]Clamp connections are specialized hyphal outgrowths unique to dikaryotic hyphae in Basidiomycota, forming at septal sites to ensure equal distribution of the two unlike nuclei during cell division. These hook-like structures develop from the penultimate hyphal compartment, fusing with the adjacent septum to create a bridge that facilitates nuclear migration and pairing. Clamp connections are absent in monokaryotic phases and are a hallmark of the dikaryotic mycelium, supporting genetic stability in this prolonged life stage.[32][33]Fungal hyphal growth follows distinct patterns, including monopodial elongation along a single primary axis with lateral branches arising from nodes, as seen in certain conidiophores and exploratory hyphae. Dichotomous branching occurs when the hyphal tip splits into two equal branches, often in response to environmental cues, promoting radial expansion of the mycelium. Developmental sequences can be acropetal, where new structures form successively toward the apex (e.g., in conidial chains), or basipetal, proceeding toward the base, influencing the orientation of spore production and colony architecture. Geniculate growth refers to the bent or knee-like curvature of hyphae or conidiophores, typically at branching points or during maturation, as observed in genera like Cladosporium. These patterns optimize nutrient capture and reproductive efficiency.[34][35][36][37]Hyphal cells exhibit nuclear variation, with homokaryotic (or monokaryotic) states featuring a single nucleus per compartment, common in haploid phases before mating. Heterokaryotic hyphae contain multiple genetically distinct nuclei within the same cytoplasm, arising from hyphal fusion or parasexuality, which promotes genetic diversity. Dikaryotic hyphae, a specialized heterokaryon in Basidiomycota and some Ascomycota, maintain precisely two unlike nuclei per cell throughout the vegetative phase, stabilized by clamp connections and enabling prolonged binucleate growth without karyogamy. These nuclear configurations underpin fungal adaptability and reproductive strategies.[38][7]
Spores and Spore Structures
In mycology, spores are microscopic reproductive units produced by fungi, serving as primary agents for dispersal and propagation. These structures vary widely in form, function, and origin, often developing from hyphal tips or specialized cells, and are essential for fungal identification due to their distinctive morphology. Spores can be unicellular or multicellular, pigmented or hyaline, and adapted for survival in diverse environments, with their characteristics providing key taxonomic clues.[39]General spore types encompass a range of non-specialized forms. An aplanospore is a non-motile, asexual spore lacking flagella, typically formed within sporangia and dispersed passively by air or water. A ballistospore refers to a forcibly discharged spore, propelled by mechanisms such as surface tension in basidiomycetes, enabling active ejection over short distances for enhanced dispersal. The chlamydospore is a thick-walled, resting spore derived from hyphal segments or conidia, designed for dormancy and resistance to adverse conditions like desiccation or cold.[39][40]Spore morphology includes diverse shapes and surface ornamentations that aid in classification. Common shapes are globose (spherical), obovoid (egg-shaped with the broader end at the base), and pyriform (pear-shaped). Ornamentations feature dictyospores, which are muriform spores with both transverse and longitudinal septa forming a net-like pattern; echinate spores covered in prominent spines; verrucose spores with wart-like projections; and spinulose spores bearing fine, small spines. These features are observed under microscopy and are critical for distinguishing genera.[41][42][41]Specialized spores are associated with particular fungal groups and reproductive modes. An ascospore is a sexual spore formed within an ascus in Ascomycota, often octad in number and variable in shape. The basidiospore develops externally on a basidium in Basidiomycota, typically four per basidium and adapted for ballistosporic discharge. A zygospore arises from the fusion of gametangia in Mucoromycota, featuring thick, spiny walls for protection. The oospore, found in Oomycota (stramenopiles often studied in mycology), is a thick-walled sexual spore produced in an oogonium following fertilization. An ustilospore is the characteristic pigmented, thick-walled spore of smut fungi (Ustilaginomycotina), aggregated in sori on host plants.[39][39][39][39][43][29]Spore wall features include structural adaptations for attachment and germination. The hilum is a basal scar marking the former attachment point of the spore to its producing structure, such as a sterigma or conidiophore. An apiculus is a small, sterigma-derived projection or knob at the spore's hilar end, prominent in basidiospores. The germ pore is a thinned, often apical or equatorial area in the spore wall through which the germ tube emerges during germination, facilitating rapid outgrowth. These elements are visible via light or electronmicroscopy and contribute to spore viability and identification.[44][41]
Reproductive Structures
Asexual Reproduction Terms
Asexual reproduction in fungi involves the production of spores or propagules through mitotic division, resulting in genetically identical offspring that facilitate dispersal and colonization without genetic recombination. This process is prevalent in many fungal groups, particularly in the Ascomycota and Mucoromycota, and encompasses various mechanisms such as conidiogenesis and sporangiogenesis.[23] Key terms describe the modes of spore formation and associated structures, emphasizing clonal propagation for rapid environmental adaptation.[45]Conidiogenesis refers to the formation of conidia, which are non-motile, asexual spores typically borne on specialized hyphae called conidiophores. These structures arise from the mycelium and elevate conidia for efficient airborne dispersal. Conidiogenesis occurs primarily through two broad modes: blastic, involving internal enlargement of a conidial initial within the conidiogenous cell, and thallic, characterized by external fragmentation or septation of pre-existing hyphal segments.[23] Blastic conidiogenesis subtypes include phialidic, where conidia are produced enteroblastically from a flask- or vase-shaped conidiogenous cell (phialide) with a fixed neck, allowing sequential basipetal formation of conidia without altering the cell's apex; and annellidic, a variant where the conidiogenous cell proliferates percurrently through a single opening, leaving ring-like scars (annellations) after successive conidia are extruded.[45] Thallic conidiogenesis, in contrast, produces arthroconidia or arthrospores by the disarticulation of hyphal compartments, often in chains, as seen in dermatophytes like Trichophyton species.[1]Conidia and related spores are collectively termed mitospores, as they arise from mitotic division rather than meiosis. These may aggregate into complex structures known as conidiomata, such as the flask-shaped pycnidium, a closed, ostiolate fruiting body lined with conidiogenous cells, or the cushion-like sporodochium, featuring a dense mat of erect conidiophores.[45] In parallel, sporangial methods involve sporangia, sac-like vesicles containing numerous spores, supported by branched sporangiophors in groups like the Mucoromycota; spores released upon maturation serve similar dispersive roles to conidia.[23]Other asexual forms include budding in yeasts, where a daughter cell emerges from the parent via localized cell wall expansion, often producing blastospores that resemble buds and enable rapid proliferation in liquid environments.[46] Additionally, gemmae are multicellular, thick-walled outgrowths from hyphae or mycelium that detach as propagules, germinating directly into new mycelia, as observed in certain soil fungi.[1] These mechanisms collectively underscore the diversity of non-sexual propagation in fungi, optimizing survival in varied ecological niches.[47]
Sexual Reproduction Terms
Sexual reproduction in fungi promotes genetic diversity through the fusion of compatible nuclei, contrasting with asexual processes by involving meiosis to produce recombinant spores. This cycle typically begins with the formation of specialized structures or hyphal fusions that bring together haploid nuclei, followed by cytoplasmic and nuclear unions, and culminates in meiotic divisions within characteristic spore-producing cells. Unlike many organisms, fungi often delay nuclear fusion after cytoplasmic merging, maintaining a prolonged dikaryotic phase that enhances adaptability.[39]In certain fungal groups, particularly Mucoromycota, sexual reproduction involves the fusion of compatible hyphae to form multinucleate gametangia that merge without distinct gametes, leading to zygospore formation. In some Chytridiomycota, sexual reproduction occurs via oogonia (female gametangia) fertilized by motile gametes produced by antheridia (male gametangia).The fusion processes central to fungal sexuality include plasmogamy, the merging of protoplasts from compatible hyphae or gametes, which establishes a heterokaryotic or dikaryotic condition without immediate nuclear fusion. This is followed by karyogamy, the union of the haploid nuclei to form a diploid zygote, typically occurring much later in specialized cells. In many Basidiomycota and some Ascomycota, somatogamy occurs instead, involving direct fusion of somatic hyphal cells without distinct gametangia, initiating the dikaryotic phase.[39][48]Fungal mating compatibility is governed by heterothallism, where self-sterile individuals require two compatible thalli or mating types for plasmogamy to promote outcrossing and genetic variation, or homothallism, in which a single self-fertile thallus can complete the sexual cycle independently, often favoring inbreeding. These systems ensure controlled recombination, with heterothallism predominant in many species to avoid homozygous disadvantage.[39]The outcomes of these fusions vary by phylum: in Ascomycota, karyogamy precedes meiosis within the ascus, a sac-like cell producing ascospores, often facilitated by crozier structures—hooked hyphal tips that divide to position nuclei for fusion and ascospore development. In Basidiomycota, meiosis occurs in the basidium, a club-shaped cell generating basidiospores on sterigmata, with dikaryotic hyphae maintained by clamp connections; variant racket cells, swollen segments in these clamps, aid in nuclear distribution during hyphal growth. In Mucoromycota, the result is a zygospore, a thick-walled, resistant diploid structure formed directly from gametangial fusion, enduring adverse conditions until germination triggers meiosis.[1][48]
Fruiting Bodies and Macrostructures
Ascomycete Structures
Ascomycete structures encompass the specialized fruiting bodies known as ascomata, which are characteristic of the phylum Ascomycota, commonly referred to as sac fungi. These structures house the asci, the sac-like cells in which sexual spores (ascospores) are produced following meiosis. Ascomata vary in morphology to facilitate spore dispersal and protection, reflecting adaptations to diverse ecological niches. The primary types include the apothecium, perithecium, and cleistothecium, each differing in openness and development.[49][50]The apothecium is an open, cup-shaped ascoma that exposes the spore-bearing layer to the air, promoting efficient ascospore discharge. Examples include the brightly colored elf cups (Sarcoscypha species), where the hymenium lines the inner surface of the cup. This structure is typical of the Pezizales order and aids in passive spore release via air currents. In contrast, the perithecium is a flask-shaped ascoma with a narrow opening called an ostiole, through which mature ascospores are forcibly ejected. This enclosed form provides protection against desiccation and predators, as seen in fungi like Neurospora. The cleistothecium, fully closed without an opening, relies on mechanical rupture or decay for spore release; it is common in powdery mildews (Erysiphales), where the spherical structure bursts irregularly.[49][50][50]Internally, ascomata feature the hymenium, a fertile layer of closely packed asci embedded in a matrix of sterile tissues. The asci themselves are elongated, sac-like cells that develop at the tips of dikaryotic hyphae after karyogamy and meiosis, typically containing eight ascospores arranged in a linear or biseriate pattern. Interspersed among the asci are paraphyses, which are sterile, filamentous hyphae that arise from the hymenial base and extend upward, aiding in spacing and potentially facilitating ascus maturation by providing structural support and nutrient transport. The collective tissue of these sterile elements, including paraphyses and other hyphae between or around the asci, is termed the hamathecium, which varies from simple filamentous to pseudoparenchymatous forms depending on the taxon.[51][52][52]In some ascomycetes, protective coverings analogous to veils occur during development, though less common than in basidiomycetes. A universal veil may envelop the immature fruiting body, leaving remnants as patches or a basal cup upon maturation, while a partial veil covers the developing asci, potentially forming an annular structure. These are observed in certain morchella-like species, enhancing protection against environmental stress. Additionally, ascostromata represent stroma-embedded ascomata, where multiple asci are immersed in a compact hyphal mass rather than forming discrete fruiting bodies; this is prevalent in plant-pathogenic groups like the Dothideomycetes. A columella, adapted as a central, pillar-like sterile axis within some ascomatal cavities, provides structural reinforcement, particularly in perithecioid forms.[53][54][11]
Basidiomycete Structures
Basidiomycete structures encompass the diverse fruiting bodies and microscopic features characteristic of the Basidiomycota, commonly known as club fungi, which produce spores on specialized club-shaped cells. These structures are primarily macroscopic in familiar forms like mushrooms but include microscopic elements essential for reproduction. The basidioma, or fruiting body, serves as the spore-producing organ, varying widely in morphology to facilitate spore dispersal in different environments.[1][55]Basidioma types include the agaric, a classic gilled mushroom with an expanded cap that exposes gills for spore release, as seen in species like those in the genus Agaricus. The bolete features a cap atop a stem but with a porous underside of tubes rather than gills, exemplified by genera such as Suillus and Boletus, where the hymenophore consists of tightly packed pores. Secotioid forms represent an intermediate morphology, with partially enclosed gills that prevent full expansion of the cap, leading to reduced spore discharge and often hypogeous tendencies, as in certain Amanita relatives. Puffballs, or gasteroid basidiomata, are globular and enclosed, containing a gleba of spore mass that releases through an apical pore upon maturity, such as in Lycoperdon species.[56][57]Key parts of these structures include the pileus, the cap-like upper portion that protects the spore-bearing surface in agarics and boletes; the stipe, a stalk that elevates the pileus for efficient dispersal; and the lamella, the thin, radiating gill plates beneath the pileus in agarics that bear reproductive cells. The hymenophore refers to the spore-producing layer, which may be lamellate (gilled), poroid (tubular), or otherwise configured depending on the basidioma type. At the microscopic level, the basidium is the club-shaped cell where meiosis occurs, typically producing four spores, while each sterigma is a narrow projection from the basidium that supports an individual basidiospore. Basidiospores form on these sterigmata and are briefly referenced here as the external spores dispersed from the hymenium.[1][55][56]Veils and accessory structures provide protection during development. The annulus is a ring-like remnant on the stipe from a partial veil that initially covers the gills. The volva forms a cup at the stipe base, derived from a universal veil enveloping the young basidioma. A cortina appears as a web-like partial veil connecting the pileus margin to the stipe, while the marginal veil similarly shields the developing hymenophore edges in immature stages.[1]Internally, the trama constitutes the fleshy tissue between lamellae or within the pileus, composed of interwoven hyphae that support the structure. Cystidia are elongated, sterile cells protruding from the hymenium, often aiding in spore discharge or species identification by their shape and ornamentation.[1][55]
Ecological and Habitat Terms
Symbioses and Interactions
In mycology, symbioses and interactions refer to the diverse partnerships fungi form with other organisms, encompassing mutualistic associations that benefit both parties through nutrient exchange and antagonistic relationships involving parasitism or predation. These interactions are fundamental to fungal ecology, influencing biodiversity, soil health, and trophic dynamics in terrestrial and aquatic environments. Mutualisms like mycorrhizae and lichens exemplify cooperative strategies where fungi provide structural support or access to soil resources in exchange for photosynthetic products, while antagonisms such as predation on nematodes highlight fungi's role as biological control agents.[58][59]Mycorrhizae represent one of the most widespread symbiotic mutualisms, occurring in approximately 80-90% of vascular plant species, where fungi colonize plant roots to enhance nutrient and water uptake. Ectomycorrhiza (ECM) form a fungal sheath, or Hartig net, around the exterior of short roots without penetrating cortical cells, typically involving basidiomycetes or ascomycetes with woody plants like pines and oaks; this sheath facilitates phosphorus and nitrogen acquisition for the host while the fungus receives carbohydrates.[60]Arbuscular mycorrhiza (AM), the most ancient and common type mediated by Glomeromycota, feature intracellular hyphae that form arbuscules—branched structures within root cortical cells—for bidirectional nutrient transfer, aiding over 70% of land plants in mineral absorption.[60] Specialized forms include ericoid mycorrhiza (ErM), where ascomycetes like those in the Leotiomycetes create intracellular coils in fine roots of Ericaceae plants (e.g., heather), enabling organic nitrogen breakdown in nutrient-poor soils, and orchid mycorrhiza (OM), a unique association with basidiomycetes or ascomycetes that supplies carbon to orchid seedlings during mycoheterotrophic phases before photosynthesis begins.[61]Lichens are stable, self-supporting symbiotic consortia between a fungal partner, the mycobiont (typically an ascomycete), and one or more photosynthetic partners, the photobiont (usually green algae like Trebouxia or cyanobacteria such as Nostoc), allowing fungi to thrive in extreme habitats by combining the mycobiont's protective thallus structure with the photobiont's fixed carbon.[62][63] Reproductive propagules in lichens include soredia, powdery clusters of photobiont cells enclosed in fungal hyphae that disperse the entire partnership via wind or water, and isidia, upright outgrowths of the thallus containing both symbionts that fragment for vegetative propagation without disrupting the lichen's integrity.[64][65]Beyond these prominent mutualisms, fungi exhibit a spectrum of other interactions, often antagonistic. Endophytes are fungi that colonize plant tissues internally without causing apparent disease, providing benefits like enhanced drought tolerance or defense against herbivores through secondary metabolite production, as seen in clavicipitaceous endophytes of grasses.[66]Fungicolous fungi grow on or within other fungi, ranging from parasitic hyperparasites that weaken hosts by nutrient depletion to saprotrophic utilizers of fungal debris, exemplified by species in the Hypocreales.[67]Lichenicolous fungi, often ascomycetes, inhabit lichen thalli as obligate parasites or commensals, deriving nutrients from the mycobiont or photobiont while sometimes altering lichen morphology.[68]Nematophagous fungi, such as those in the Orbiliaceae or Monacrosporium, actively trap and digest nematodes using adhesive hyphae, constricting rings, or toxic compounds, serving as natural biocontrol agents against plant-parasitic nematodes in soil ecosystems.[69][70] A key morphological adaptation in many parasitic and symbiotic fungi is the haustorium, a specialized hyphal extension that penetrates host cells to absorb nutrients while enclosed by an extrahaustorial matrix, essential for biotrophic pathogens like rust fungi during compatible interactions.[71]
Habitat Preferences
Fungi exhibit diverse habitat preferences that reflect their ecological roles and adaptations to specific environmental conditions and substrates. These preferences are often described through terms that denote trophic modes, substrate affinities, and growth patterns, enabling mycologists to classify fungal distributions accurately. Such terminology highlights how fungi colonize various niches, from decaying organic matter to extreme environments, without relying on biotic interactions like symbiosis.Trophic modes primarily categorize fungi based on their nutritional strategies in habitats. Saprobe refers to fungi that derive nutrients from decaying organic matter, acting as decomposers in ecosystems by breaking down dead plant and animal material.[27]Lignicolous fungi are those that grow on or in wood, often specializing in the degradation of lignin-rich substrates like fallen logs or timber. Coprophilous species colonize dung, playing a key role in nutrientrecycling in herbivore-dominated environments. Foliicolous fungi thrive on leaf surfaces, typically as epiphytes or initial decomposers in forest canopies.Substrate-based terms further specify fungal affinities for abiotic or semi-abiotic environments. Aquatic fungi inhabit freshwater or marine ecosystems, where they contribute to organic matter decomposition in submerged habitats. Aero-aquatic fungi are adapted to interfaces between air and water, forming specialized structures like bubble-forming conidia to facilitate spore dispersal in moist, aerial conditions. Geophilous fungi grow in or on soil, often as primary colonizers of terrestrial substrates. Saxicolous species adhere to rock surfaces, enduring harsh, exposed conditions through endolithic or epilithic growth. Corticolous fungi establish on tree bark, exploiting the nutrient-rich interface between living wood and atmosphere.Specialized habitat terms denote narrower adaptations. Algicolous fungi grow on or within algae, frequently in aquatic or moist terrestrial settings where they may act as parasites or commensals. Fructicolous species develop on the fruiting bodies of other fungi, utilizing these ephemeral resources for sporulation. Ramicolous fungi colonize twigs or small branches, often in litter layers or arboreal niches. Xerophilic fungi are tolerant of low water activity, surviving in arid environments like desiccated soils or stored grains by producing protective spores or sclerotia.Growth patterns describe the morphological expression of habitat colonization. Effuse growth involves thin, spreading mycelium that forms expansive, non-discrete patches on substrates. Crustose forms create a compact, crust-like thallus, typically on stable surfaces like wood or rock. Fairy ring patterns arise from radial mycelial expansion in soil, manifesting as circular fruiting bodies in grasslands due to nutrient depletion at the mycelium's advancing edge.[72]
Pathogenicity and Human Impact
Fungal Diseases
Fungal diseases, or mycoses, encompass a wide range of infections caused by pathogenic fungi that affect plants, animals, and humans, often leading to significant economic, ecological, and health impacts. These pathogens exploit host vulnerabilities through direct tissue invasion, spore dissemination, or environmental persistence, with symptoms varying from localized lesions to systemic failures. In plants, fungal diseases frequently target crops and forests, causing yield losses and structural weakening, while in animals and humans, they range from superficial skin conditions to life-threatening systemic infections. Emerging fungal threats, such as those driving amphibian declines, highlight the evolving role of fungi in biodiversity loss.[73]In plants, rust diseases are caused by obligate parasitic fungi in the subphylum Pucciniomycotina (order Pucciniales), which produce characteristic rust-colored pustules containing urediniospores on leaves, stems, and fruits, leading to chlorosis, necrosis, and reduced photosynthesis. These pathogens often require alternate hosts to complete their complex life cycles and can devastate cereal crops like wheat and barley.[74] Smut diseases, induced by biotrophic fungi in the subphylum Ustilaginomycotina, replace host tissues—particularly flowers, grains, or stems—with masses of teliospores that appear as black or sooty powders, severely compromising seed production in grasses and cereals such as corn and sorghum. Potato blight, while caused by the oomycetePhytophthora infestans (not a true fungus but often discussed in fungal pathology contexts), manifests as dark lesions on foliage and tubers, resulting in rapid plant collapse and historical famines through sporangia dispersal in cool, moist conditions.[75] Ergot disease, produced by ascomycete fungi in the genus Claviceps (primarily C. purpurea), replaces grain kernels with hard, dark sclerotia that contain toxic alkaloids, affecting rye and other grasses and posing risks to livestock and humans via contaminated feed.Wood-decay fungi, primarily basidiomycetes, act as pathogens in trees by invading wounded or stressed wood, causing structural weakening that predisposes trees to breakage. White-rot fungi degrade both lignin and cellulose, resulting in a spongy, bleached appearance of the wood and affecting hardwoods like oaks and maples, with examples including species of Ganoderma and Trametes. In contrast, brown-rot fungi selectively break down cellulose and hemicellulose while modifying lignin, producing dry, cubical, and brittle wood fragments, commonly impacting conifers such as pines, as seen with Laetiporus species.[76][77]In humans and animals, mycosis refers to any disease resulting from fungal infection, ranging from superficial to deep-seated invasions that can disseminate via bloodstream or inhalation of spores. Dermatophytosis, also known as ringworm or tinea, is a superficial infection of the skin, hair, or nails caused by dermatophytes—keratinophilic fungi such as Trichophyton, Microsporum, and Epidermophyton species—that produce annular lesions with scaling and itching, thriving in warm, moist environments.[78][79] Mycetoma is a chronic subcutaneous infection characterized by swelling, sinus tracts, and grain-like discharge, often initiated by traumatic inoculation of fungi like Madurella species in soil-contaminated wounds, predominantly affecting extremities in tropical regions.[80] Piedra involves fungal colonization of hair shafts, forming hard nodules: white piedra by Trichosporon species on scalp hairs and black piedra by Piedraia hortae on facial hairs, leading to breakage without systemic spread. Endothrix refers to a pattern of hair infection where fungal spores fill the hair shaft interior without external sheath formation, typically seen in dermatophytoses like tinea capitis caused by Trichophyton tonsurans.[81] Dimorphic pathogens, such as Histoplasma capsulatum, exhibit temperature-dependent morphology—mold at environmental temperatures and yeast in host tissues—causing pulmonary and disseminated infections via inhalation of conidia from soil enriched with bird or bat guano, with severe cases in immunocompromised individuals. An emerging human fungal pathogen is Candida auris, a multidrug-resistant yeast causing invasive infections, particularly in healthcare settings, with global outbreaks reported since 2009 and increasing incidence as of 2025.[82]An emerging fungal disease, chytridiomycosis, is caused by the chytrid fungus Batrachochytrium dendrobatidis and infects amphibian skin, disrupting electrolyte balance and leading to cardiac arrest, contributing to the decline or extinction of at least 501 species worldwide, with over 90 presumed extinct, particularly in tropical regions.[83][84]
Mycotoxins and Uses
Mycotoxins are secondary metabolites produced by certain fungi that can adversely affect human and animal health upon ingestion, inhalation, or dermal contact. These toxic compounds are often generated by species in genera such as Aspergillus, Penicillium, and Fusarium, contaminating food crops like grains, nuts, and fruits under favorable environmental conditions.[85] Aflatoxins, produced primarily by Aspergillus flavus and A. parasiticus, are among the most potent mycotoxins, causing acute liver damage and chronic effects like hepatocellular carcinoma; they contaminate crops such as peanuts, corn, and rice.[86] Ergot alkaloids, synthesized by Claviceps purpurea on rye and other cereals, induce ergotism, manifesting as hallucinations, convulsions, and vasoconstriction; these compounds have also served as precursors for pharmaceuticals like ergotamine.[87] Patulin, produced by Penicillium expansum during apple rot, poses risks in fruit juices and ciders, causing gastrointestinal distress and potential neurotoxicity at high exposures.[85]Despite their hazards, fungi yield numerous beneficial compounds and processes exploited by humans. Yeasts, particularly Saccharomyces cerevisiae, drive alcoholic fermentation in bread, beer, and wine production by converting sugars into ethanol and carbon dioxide, enabling leavening and flavor development essential to global cuisines.[88] Medicinal applications include penicillin, an antibiotic derived from Penicillium chrysogenum (formerly P. notatum), discovered serendipitously by Alexander Fleming in 1928 when mold inhibited bacterial growth in culture; mass production revolutionized treatment of bacterial infections during World War II.[89] Trehalose, a non-reducing disaccharide accumulated in fungal spores for stress protection, finds industrial uses as a stabilizer in foods, pharmaceuticals, and cosmetics due to its ability to preserve proteins and cells under desiccation or freezing.[90]Key terms in this domain encompass both protective and cultural dimensions. A fungicide is a chemical agent designed to inhibit or kill fungi, widely applied in agriculture to control crop pathogens like Botrytis cinerea.[91]Ethnomycology examines human-fungal interactions across cultures, including traditional uses of mushrooms for food, medicine, and rituals, such as psychoactive species in indigenous practices.[92]Noble rot refers to a beneficial infection by Botrytis cinerea on grapes under humid, warm conditions, concentrating sugars and imparting complex flavors to dessert wines like Sauternes.[93] Human impacts involve fungivory, where animals or insects consume fungi as a primary diet, influencing spore dispersal; in humans, this extends to mycetism, or poisoning from ingesting toxic mushrooms, often due to amatoxins or other mycotoxins causing severe gastrointestinal and hepatic symptoms.[94]
Advanced and Modern Concepts
Genetics and Molecular Biology
Fungal genetics encompasses the study of inheritance, variation, and molecular mechanisms that underpin fungal biology, including nuclear division processes unique to these organisms. Mitosis in filamentous fungi involves asynchronous nuclear divisions within shared cytoplasm, allowing for rapid hyphal extension without cytokinesis, a process that differs from the synchronous division seen in many higher eukaryotes.[95] Meiosis in fungi, such as in Coprinus cinereus, is regulated by environmental cues like light-dark cycles, which trigger karyogamy and progression through meiotic stages to produce basidiospores.[96] In basal fungal lineages like chytrids, the kinetosome—equivalent to the basal body—organizes the flagellar apparatus in zoospores, featuring a distinctive ultrastructure with microtubular singlets that supports motility in aquatic environments.[97]Advancements in fungal genomics have revealed extensive diversity through large-scale sequencing initiatives, such as the 1000 Fungal Genomes Project, which has sequenced representatives from over 500 fungal families (as of 2023) to elucidate evolutionary patterns and metabolic capabilities.[98][99] These efforts highlight genome architectures adapted to ecological niches, including compact genomes in yeasts and expansive ones in filamentous species with repetitive elements. Karyotype variations are prevalent in fungi, particularly among plant pathogens, where chromosomal rearrangements like translocations and duplications contribute to adaptability and virulence without altering gene content significantly.[100] For instance, intraspecific polymorphisms in chromosome number and size enable rapid evolution in response to host defenses.Modern molecular tools like CRISPR-Cas9 have revolutionized fungal gene editing, enabling precise modifications in filamentous species such as Aspergillus and Neurospora to study gene function and enhance traits like metabolite production.[101] Applications include targeted disruptions of pathogenicity genes and insertion of reporters, overcoming previous limitations in homologous recombination efficiency. Anastomosis, or hyphal fusion, facilitates vegetative gene transfer between compatible hyphae, promoting resource sharing and genetic heterogeneity within mycelia.[102] This process often leads to heterokaryons—multinucleate cells harboring genetically distinct nuclei—creating somatic mosaics that can mask deleterious mutations or drive adaptive evolution.[103] In fungal pathogens, physiological races denote strains differentiated by virulence on specific host genotypes, arising from genetic variations that alter effector profiles without taxonomic distinction.[104]The mycovirome refers to the collective viral community infecting fungi, comprising diverse RNA and DNA viruses that can attenuate host virulence or influence population dynamics.[105] Gene clusters for secondary metabolites, often spanning 10-100 kb and containing 5-20 biosynthetic genes, are key to fungal chemical ecology, regulated by chromatin remodeling and environmental signals to produce antibiotics, toxins, and signaling molecules.[106] These clusters exhibit modular organization, with core enzymes like polyketide synthases driving structural diversity essential for survival and biotechnological applications.
Ecological Networks and Mycobiome
In fungal ecology, ecological networks refer to the interconnected systems formed by mycelial structures that facilitate resource exchange, signaling, and interactions among plants, microbes, and other organisms within ecosystems. These networks, often spanning large areas, enable the transfer of nutrients, water, and chemical signals, influencing community dynamics and resilience. A prominent example is the "wood wide web," a term coined to describe the mycorrhizal networks linking tree roots through shared fungal hyphae, allowing bidirectional carbon and nutrient flow between plants, although the extent and adaptive significance of these interactions remain subjects of scientific debate.[107] Seminal research demonstrated net carbon transfer from mature trees to seedlings via ectomycorrhizal fungi in forest understories, highlighting how these networks support kin recognition and resource partitioning among species.[108]Fungal highways, exemplified by rhizomorphs—aggregates of hyphae forming cord-like structures in basidiomycetes such as Armillaria—serve as efficient transport conduits for water, nutrients, and organic compounds across substrates. These structures, up to several meters long, create hydraulic pathways that redistribute resources in heterogeneous environments like forest soils, enhancing fungal foraging efficiency and connectivity in nutrient-poor areas. Studies have shown rhizomorphs can bridge dry and moist soil zones, facilitating moisture transport to support mycelial growth and host plants during drought.The mycobiome encompasses the assemblage of fungal taxa inhabiting a specific host or environmental niche, interacting with bacterial microbiota to shape ecosystem processes such as decomposition and nutrientcycling. In hosts like plants or animals, the mycobiome modulates immune responses and metabolic functions, while in soils or aquatic systems, it contributes to organic matter breakdown and pollutant remediation. Broader microbiota includes these fungal components alongside bacteria, archaea, and protists, forming holobionts that enhance host adaptability; for instance, plant mycobiomes can alleviate stress from pathogens through competitive exclusion. The term mycobiome was formalized in studies of human-associated fungi, but its application extends to environmental contexts where fungal diversity influences community stability.[109]Key terms in fungal community ecology include autochthonous fungi, which are indigenous species native to and evolved within a particular habitat, contrasting with introduced allochthonous forms; these native fungi often dominate stable microbial assemblages in soils or sediments, driving localized nutrient cycles. Ruderal fungi, akin to pioneer species, are opportunistic colonizers of disturbed or barren substrates, such as post-fire soils or decaying wood, exhibiting high reproductive rates and stress tolerance to initiate succession. For example, ruderal basidiomycetes rapidly degrade simple lignocellulose, paving the way for more competitive successors. Senescence describes the progressive decline and eventual cessation of growth in fungal colonies, often linked to mitochondrial dysfunction and oxidative stress, limiting colony lifespan in aging mycelia. This phenomenon is well-documented in model organisms like Neurospora, where senescing cultures exhibit reduced vigor due to accumulated cellular damage.[110]Contemporary research has unveiled "fungal dark matter," referring to the vast uncultured fungal diversity detected through environmental metagenomics, representing up to 90% of estimated fungal species that evade traditional culturing methods. This hidden fraction includes basal lineages with novel ecologies, challenging phylogenetic frameworks and underscoring gaps in our understanding of global fungal roles in carbon sequestration and bioremediation. In aquatic ecosystems, Rozellomycota (previously classified as Rozellida)—a phylum of early-diverging, zoosporic fungi—play pivotal roles as endoparasites of algae, oomycetes, and other microbes, regulating population dynamics and nutrient release in freshwater and marine habitats. Their thin-walled zoospores and intracellular lifestyle enable efficient dispersal and host exploitation, contributing to microbial food web stability.Climate change profoundly alters fungal distributions by shifting temperature and precipitation regimes, favoring thermotolerant species while contracting ranges of cold-adapted ectomycorrhizae in boreal forests. Meta-analyses reveal that global fungal biogeography correlates strongly with climatic variables, with projections indicating poleward migrations and potential losses in tropical diversity, disrupting symbiotic networks and decomposition rates. These shifts exacerbate ecosystem vulnerabilities, as seen in reduced mycorrhizal colonization under drought, amplifying feedbacks on plant productivity and soil health.[111]




















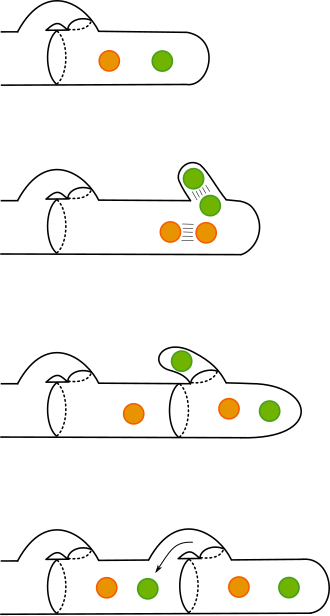



















.JPG)









_op_dode_tak_van_een_eik_15-01-2021._(actm.)_02.jpg)













































.jpg)










