Community hub
Recent from talks
Knowledge base stats:
Talk channels stats:
Members stats:
Neuroimmune system
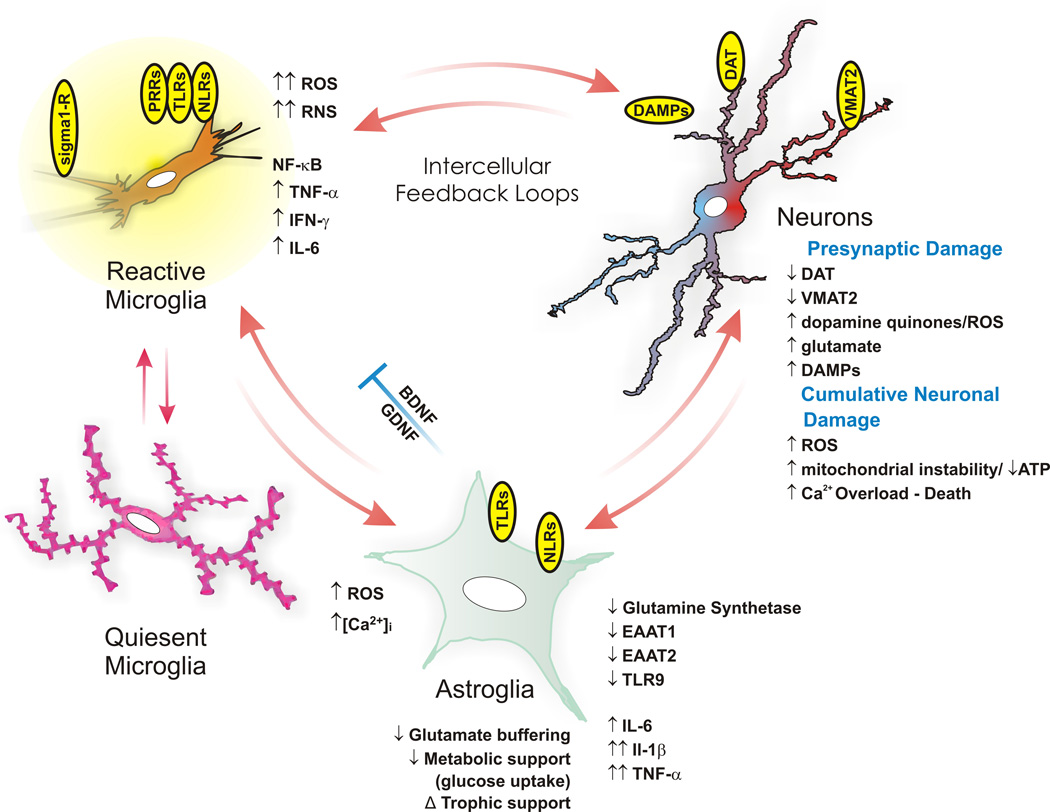
The neuroimmune system is a system of structures and processes involving the biochemical and electrophysiological interactions between the nervous system and immune system which protect neurons from pathogens. It serves to protect neurons against disease by maintaining selectively permeable barriers (e.g., the blood–brain barrier and blood–cerebrospinal fluid barrier), mediating neuroinflammation and wound healing in damaged neurons, and mobilizing host defenses against pathogens.
The neuroimmune system and peripheral immune system are structurally distinct. Unlike the peripheral system, the neuroimmune system is composed primarily of glial cells; among all the hematopoietic cells of the immune system, only mast cells are normally present in the neuroimmune system. However, during a neuroimmune response, certain peripheral immune cells are able to cross various blood or fluid–brain barriers in order to respond to pathogens that have entered the brain. For example, there is evidence that following injury macrophages and T cells of the immune system migrate into the spinal cord. Production of immune cells of the complement system have also been documented as being created directly in the central nervous system.
The key cellular components of the neuroimmune system are glial cells, including astrocytes, microglia, and oligodendrocytes. Unlike other hematopoietic cells of the peripheral immune system, mast cells naturally occur in the brain where they mediate interactions between gut microbes, the immune system, and the central nervous system as part of the microbiota–gut–brain axis.
G protein-coupled receptors that are present in both CNS and immune cell types and which are responsible for a neuroimmune signaling process include:
Neuroimmunity is additionally mediated by the enteric nervous system, namely the interactions of enteric neurons and glial cells. These engage with enteroendocrine cells and local macrophages, sensing signals from the gut lumen, including those from the microbiota. These signals prompt local immune responses and transmit to the CNS through humoral and neural pathways. Interleukins and signals from immune cells can access the hypothalamus via the neurovascular unit or circumventricular organs.
The neuro-immune system, and study of, comprises an understanding of the immune and neurological systems and the cross-regulatory impacts of their functions. Cytokines regulate immune responses, possibly through activation of the hypothalamic-pituitary-adrenal (HPA) axis.[medical citation needed] Cytokines have also been implicated in the coordination between the nervous and immune systems. Instances of cytokine binding to neural receptors have been documented between the cytokine releasing immune cell IL-1 β and the neural receptor IL-1R. This binding results in an electrical impulse that creates the sensation of pain. Growing evidence suggests that auto-immune T-cells are involved in neurogenesis. Studies have shown that during times of adaptive immune system response, hippocampal neurogenesis is increased, and conversely that auto-immune T-cells and microglia are important for neurogenesis (and so memory and learning) in healthy adults.
The neuroimmune system uses complementary processes of both sensory neurons and immune cells to detect and respond to noxious or harmful stimuli. For example, invading bacteria may simultaneously activate inflammasomes, which process interleukins (IL-1 β), and depolarize sensory neurons through the secretion of hemolysins. Hemolysins create pores causing a depolarizing release of potassium ions from inside the eukaryotic cell and an influx of calcium ions. Together this results in an action potential in sensory neurons and the activation of inflammasomes.
Injury and necrosis also cause a neuroimmune response. The release of adenosine triphosphate (ATP) from damaged cells binds to and activates both P2X7 receptors on macrophages of the immune system, and P2X3 receptors of nociceptors of the nervous system. This causes the combined response of both a resulting action potential due to the depolarization created by the influx of calcium and potassium ions, and the activation of inflammasomes. The produced action potential is also responsible for the sensation of pain, and the immune system produces IL-1 β as a result of the ATP P2X7 receptor binding.
Hub AI
Neuroimmune system AI simulator
(@Neuroimmune system_simulator)
Neuroimmune system
The neuroimmune system is a system of structures and processes involving the biochemical and electrophysiological interactions between the nervous system and immune system which protect neurons from pathogens. It serves to protect neurons against disease by maintaining selectively permeable barriers (e.g., the blood–brain barrier and blood–cerebrospinal fluid barrier), mediating neuroinflammation and wound healing in damaged neurons, and mobilizing host defenses against pathogens.
The neuroimmune system and peripheral immune system are structurally distinct. Unlike the peripheral system, the neuroimmune system is composed primarily of glial cells; among all the hematopoietic cells of the immune system, only mast cells are normally present in the neuroimmune system. However, during a neuroimmune response, certain peripheral immune cells are able to cross various blood or fluid–brain barriers in order to respond to pathogens that have entered the brain. For example, there is evidence that following injury macrophages and T cells of the immune system migrate into the spinal cord. Production of immune cells of the complement system have also been documented as being created directly in the central nervous system.
The key cellular components of the neuroimmune system are glial cells, including astrocytes, microglia, and oligodendrocytes. Unlike other hematopoietic cells of the peripheral immune system, mast cells naturally occur in the brain where they mediate interactions between gut microbes, the immune system, and the central nervous system as part of the microbiota–gut–brain axis.
G protein-coupled receptors that are present in both CNS and immune cell types and which are responsible for a neuroimmune signaling process include:
Neuroimmunity is additionally mediated by the enteric nervous system, namely the interactions of enteric neurons and glial cells. These engage with enteroendocrine cells and local macrophages, sensing signals from the gut lumen, including those from the microbiota. These signals prompt local immune responses and transmit to the CNS through humoral and neural pathways. Interleukins and signals from immune cells can access the hypothalamus via the neurovascular unit or circumventricular organs.
The neuro-immune system, and study of, comprises an understanding of the immune and neurological systems and the cross-regulatory impacts of their functions. Cytokines regulate immune responses, possibly through activation of the hypothalamic-pituitary-adrenal (HPA) axis.[medical citation needed] Cytokines have also been implicated in the coordination between the nervous and immune systems. Instances of cytokine binding to neural receptors have been documented between the cytokine releasing immune cell IL-1 β and the neural receptor IL-1R. This binding results in an electrical impulse that creates the sensation of pain. Growing evidence suggests that auto-immune T-cells are involved in neurogenesis. Studies have shown that during times of adaptive immune system response, hippocampal neurogenesis is increased, and conversely that auto-immune T-cells and microglia are important for neurogenesis (and so memory and learning) in healthy adults.
The neuroimmune system uses complementary processes of both sensory neurons and immune cells to detect and respond to noxious or harmful stimuli. For example, invading bacteria may simultaneously activate inflammasomes, which process interleukins (IL-1 β), and depolarize sensory neurons through the secretion of hemolysins. Hemolysins create pores causing a depolarizing release of potassium ions from inside the eukaryotic cell and an influx of calcium ions. Together this results in an action potential in sensory neurons and the activation of inflammasomes.
Injury and necrosis also cause a neuroimmune response. The release of adenosine triphosphate (ATP) from damaged cells binds to and activates both P2X7 receptors on macrophages of the immune system, and P2X3 receptors of nociceptors of the nervous system. This causes the combined response of both a resulting action potential due to the depolarization created by the influx of calcium and potassium ions, and the activation of inflammasomes. The produced action potential is also responsible for the sensation of pain, and the immune system produces IL-1 β as a result of the ATP P2X7 receptor binding.