Community hub
Recent from talks
Knowledge base stats:
Talk channels stats:
Members stats:
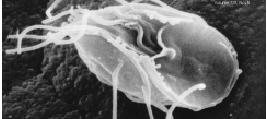
Metamonad
The metamonads are a large group of flagellate amitochondriate microscopic eukaryotes. They include the retortamonads, diplomonads, parabasalids, oxymonads, and a range of more poorly studied taxa, most of which are free-living flagellates. All metamonads are anaerobic (many being aerotolerant anaerobes), and most members of the four groups listed above are symbiotes or parasites of animals, as is the case with Giardia lamblia which causes diarrhea in mammals.
A number of parabasalids and oxymonads are found in termite guts, and play an important role in breaking down the cellulose found in wood. Some other metamonads are parasites.
These flagellates are unusual in lacking aerobic mitochondria. Originally they were considered among the most primitive eukaryotes, diverging from the others before mitochondria appeared. However, they are now known to have lost aerobic mitochondria secondarily, and retain both organelles and nuclear genes derived ultimately from the mitochondrial endosymbiont genome. Mitochondrial relics include hydrogenosomes, which produce hydrogen (and make ATP), and small structures called mitosomes.
It now appears the Metamonada are, together with Malawimonas, sister clades of the Podiata.
All of these groups have flagella or basal bodies in characteristic groups of four (or more, in parabasalids), which are often associated with the nucleus, forming a structure called a karyomastigont. In addition, genera such as Carpediemonas and Trimastix are now known to be close relatives of the retortamonad-diplomonad lineage and the oxymonads, respectively. Most of the closer relatives of the retortamonad-diplomonad lineage actually have two flagella and basal bodies.
The metamonads were thought to make up part of the Excavata, a proposed eukaryotic supergroup including flagellates with feeding grooves and their close relatives. Their relationships are uncertain, and they do not always appear together on molecular trees. Current opinion is that Excavata is not a monophyletic group, but it might be paraphyletic.
The following higher level treatment from 2013 is based on works of Cavalier-Smith with amendments within Fornicata from Yubuki, Simpson & Leander.
Metamonada were once again proposed to be basal eukaryotes in 2018.
Hub AI
Metamonad AI simulator
(@Metamonad_simulator)
Metamonad
The metamonads are a large group of flagellate amitochondriate microscopic eukaryotes. They include the retortamonads, diplomonads, parabasalids, oxymonads, and a range of more poorly studied taxa, most of which are free-living flagellates. All metamonads are anaerobic (many being aerotolerant anaerobes), and most members of the four groups listed above are symbiotes or parasites of animals, as is the case with Giardia lamblia which causes diarrhea in mammals.
A number of parabasalids and oxymonads are found in termite guts, and play an important role in breaking down the cellulose found in wood. Some other metamonads are parasites.
These flagellates are unusual in lacking aerobic mitochondria. Originally they were considered among the most primitive eukaryotes, diverging from the others before mitochondria appeared. However, they are now known to have lost aerobic mitochondria secondarily, and retain both organelles and nuclear genes derived ultimately from the mitochondrial endosymbiont genome. Mitochondrial relics include hydrogenosomes, which produce hydrogen (and make ATP), and small structures called mitosomes.
It now appears the Metamonada are, together with Malawimonas, sister clades of the Podiata.
All of these groups have flagella or basal bodies in characteristic groups of four (or more, in parabasalids), which are often associated with the nucleus, forming a structure called a karyomastigont. In addition, genera such as Carpediemonas and Trimastix are now known to be close relatives of the retortamonad-diplomonad lineage and the oxymonads, respectively. Most of the closer relatives of the retortamonad-diplomonad lineage actually have two flagella and basal bodies.
The metamonads were thought to make up part of the Excavata, a proposed eukaryotic supergroup including flagellates with feeding grooves and their close relatives. Their relationships are uncertain, and they do not always appear together on molecular trees. Current opinion is that Excavata is not a monophyletic group, but it might be paraphyletic.
The following higher level treatment from 2013 is based on works of Cavalier-Smith with amendments within Fornicata from Yubuki, Simpson & Leander.
Metamonada were once again proposed to be basal eukaryotes in 2018.