Recent from talks
Lessepsian migration
Knowledge base stats:
Talk channels stats:
Members stats:
Lessepsian migration
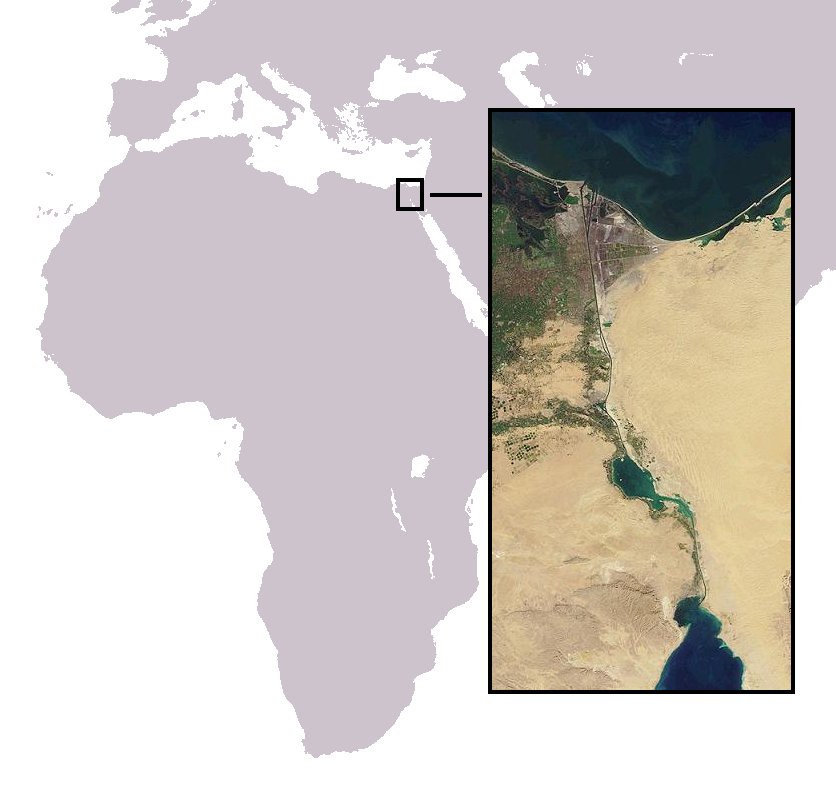
The Lessepsian migration (or Erythrean invasion) is the migration of marine species along the Suez Canal, usually from the Red Sea to the Mediterranean Sea, and more rarely in the opposite direction. When the canal was completed in 1869, fish, crustaceans, mollusks, and other marine animals and plants were exposed to an artificial passage between the two naturally separate bodies of water, and cross-contamination was made possible between formerly isolated ecosystems. The phenomenon is still occurring today. It is named after Ferdinand de Lesseps, the French diplomat in charge of the canal's construction. The term was coined by Francis Dov Por in his 1978 book.
The migration of invasive species through the Suez Canal from the Indo-Pacific region has been facilitated by many factors, both abiotic and anthropogenic, and presents significant implications for the ecological health and economic stability of the contaminated areas; of particular concern is the fisheries industry in the Eastern Mediterranean. Despite these threats, the phenomenon has allowed scientists to study an invasive event on a large scale in a short period of time, which usually takes hundreds of years in natural conditions.
The opening of the Suez Canal created the first saltwater passage between the Mediterranean Sea and the Red Sea. Constructed in 1869 to provide a more direct trade route from Europe to India and the Far East, the canal is 162.5 km (101.0 mi) long, with a depth of 10–15 m (33–49 ft) and a width varying between 200 and 300 m (660 and 980 ft).
Because the surface of the Red Sea is slightly higher in elevation than the eastern Mediterranean, the canal serves as a tidal strait by which Red Sea water pours into the Mediterranean. The Bitter Lakes, which are natural hypersaline lakes that form part of the canal, blocked the migration of Red Sea species into the Mediterranean for many decades, but as the salinity of the lakes gradually equalized with that of the Red Sea, the barrier to migration was removed, and plants and animals from the Red Sea began to colonize the eastern Mediterranean. The Red Sea, an extension of the Indian Ocean, is generally saltier and less nutrient-rich than the Mediterranean, an extension of the Atlantic Ocean, so Red Sea species, able to tolerate harsh environments, have advantages over Atlantic species in the conditions of the Eastern Mediterranean. Accordingly, most migrations between the two bodies of water are invasions of Red Sea species into the Mediterranean, and relatively few migrations occur in the opposite direction. The construction of the Aswan High Dam across the Nile River in the 1960s reduced the inflow of fresh water and nutrient-rich silt from the Nile into the eastern Mediterranean, making conditions in the eastern Mediterranean even more like those of the Red Sea, thereby increasing the impact of the invasions and facilitating the occurrence of new ones.
The Red Sea is a profusely abundant tropical marine environment sharing species in common with the eastern Indo-Pacific region, while the Mediterranean is a temperate sea with much lower productivity; the two ecosystems are extremely different in terms of structure and ecology. The Suez Canal quickly became the main pathway for the introduction of invasive species into the Eastern Mediterranean, having zoogeographic and ecological consequences far beyond what the designers foresaw. The Lessepsian migration includes hundreds of Red Sea and Indo-Pacific species that have colonized and established themselves in the Eastern Mediterranean system, causing biogeographic changes without precedent in human memory. The trend is accelerating: to take just the fish, a long-term cross-basin survey engaged by the Mediterranean Science Commission recently documented that in the first twenty years of our century more fish species from the Indo-Pacific Ocean had reached the Mediterranean than during the entire 20th century.
As of about 2010, over 1,000 species — both vertebrates and invertebrates — native to the Red Sea had been identified in the Mediterranean Sea. It was thought that many others were as yet unidentified. From there they have spread even further afield, supplying 95% of Indo-Pacific species that have reached the Ponto-Caspian seas and increasingly rapidly. In the late 20th and early 21st centuries, the Egyptian government announced its intentions to deepen and widen the canal, which raised concerns from marine biologists, fearing this would facilitate the crossing of the canal for additional species, accelerating the invasion of Red Sea species into the Mediterranean.
A wide-ranging species in the eastern Atlantic and Mediterranean, the meagre Argyrosomus regius is a species indigenous to the Eastern Mediterranean and was one of the most common commercial fish in the Levant. It has since disappeared from local catches, while the narrow-barred Spanish mackerel Scomberomorus commerson, a known Lessepsian migrant, has dramatically increased in population. Studies performed on this occurrence conclude that, due to similar life histories and diets, this may be an example of an invasive migrant outcompeting a native species and occupying its niche.
Eight species of invasive prawns from the Erythraean Sea have been recorded in the Eastern Mediterranean. These prawns are considered highly prized in Levantine fisheries, and compose most of the prawn catch off the Mediterranean coast of Egypt, being 6% of total Egyptian landings. This high abundance of invasive prawns has led to the decline of a native penaeid prawn, Melicertus kerathurus, which supported a commercial Israeli fishery throughout the 1950s. Due to outcompetition and its habitat being overrun by these migrants, this native species has since disappeared, with resultant detrimental impacts on the commercial fishery.
Hub AI
Lessepsian migration AI simulator
(@Lessepsian migration_simulator)
Lessepsian migration
The Lessepsian migration (or Erythrean invasion) is the migration of marine species along the Suez Canal, usually from the Red Sea to the Mediterranean Sea, and more rarely in the opposite direction. When the canal was completed in 1869, fish, crustaceans, mollusks, and other marine animals and plants were exposed to an artificial passage between the two naturally separate bodies of water, and cross-contamination was made possible between formerly isolated ecosystems. The phenomenon is still occurring today. It is named after Ferdinand de Lesseps, the French diplomat in charge of the canal's construction. The term was coined by Francis Dov Por in his 1978 book.
The migration of invasive species through the Suez Canal from the Indo-Pacific region has been facilitated by many factors, both abiotic and anthropogenic, and presents significant implications for the ecological health and economic stability of the contaminated areas; of particular concern is the fisheries industry in the Eastern Mediterranean. Despite these threats, the phenomenon has allowed scientists to study an invasive event on a large scale in a short period of time, which usually takes hundreds of years in natural conditions.
The opening of the Suez Canal created the first saltwater passage between the Mediterranean Sea and the Red Sea. Constructed in 1869 to provide a more direct trade route from Europe to India and the Far East, the canal is 162.5 km (101.0 mi) long, with a depth of 10–15 m (33–49 ft) and a width varying between 200 and 300 m (660 and 980 ft).
Because the surface of the Red Sea is slightly higher in elevation than the eastern Mediterranean, the canal serves as a tidal strait by which Red Sea water pours into the Mediterranean. The Bitter Lakes, which are natural hypersaline lakes that form part of the canal, blocked the migration of Red Sea species into the Mediterranean for many decades, but as the salinity of the lakes gradually equalized with that of the Red Sea, the barrier to migration was removed, and plants and animals from the Red Sea began to colonize the eastern Mediterranean. The Red Sea, an extension of the Indian Ocean, is generally saltier and less nutrient-rich than the Mediterranean, an extension of the Atlantic Ocean, so Red Sea species, able to tolerate harsh environments, have advantages over Atlantic species in the conditions of the Eastern Mediterranean. Accordingly, most migrations between the two bodies of water are invasions of Red Sea species into the Mediterranean, and relatively few migrations occur in the opposite direction. The construction of the Aswan High Dam across the Nile River in the 1960s reduced the inflow of fresh water and nutrient-rich silt from the Nile into the eastern Mediterranean, making conditions in the eastern Mediterranean even more like those of the Red Sea, thereby increasing the impact of the invasions and facilitating the occurrence of new ones.
The Red Sea is a profusely abundant tropical marine environment sharing species in common with the eastern Indo-Pacific region, while the Mediterranean is a temperate sea with much lower productivity; the two ecosystems are extremely different in terms of structure and ecology. The Suez Canal quickly became the main pathway for the introduction of invasive species into the Eastern Mediterranean, having zoogeographic and ecological consequences far beyond what the designers foresaw. The Lessepsian migration includes hundreds of Red Sea and Indo-Pacific species that have colonized and established themselves in the Eastern Mediterranean system, causing biogeographic changes without precedent in human memory. The trend is accelerating: to take just the fish, a long-term cross-basin survey engaged by the Mediterranean Science Commission recently documented that in the first twenty years of our century more fish species from the Indo-Pacific Ocean had reached the Mediterranean than during the entire 20th century.
As of about 2010, over 1,000 species — both vertebrates and invertebrates — native to the Red Sea had been identified in the Mediterranean Sea. It was thought that many others were as yet unidentified. From there they have spread even further afield, supplying 95% of Indo-Pacific species that have reached the Ponto-Caspian seas and increasingly rapidly. In the late 20th and early 21st centuries, the Egyptian government announced its intentions to deepen and widen the canal, which raised concerns from marine biologists, fearing this would facilitate the crossing of the canal for additional species, accelerating the invasion of Red Sea species into the Mediterranean.
A wide-ranging species in the eastern Atlantic and Mediterranean, the meagre Argyrosomus regius is a species indigenous to the Eastern Mediterranean and was one of the most common commercial fish in the Levant. It has since disappeared from local catches, while the narrow-barred Spanish mackerel Scomberomorus commerson, a known Lessepsian migrant, has dramatically increased in population. Studies performed on this occurrence conclude that, due to similar life histories and diets, this may be an example of an invasive migrant outcompeting a native species and occupying its niche.
Eight species of invasive prawns from the Erythraean Sea have been recorded in the Eastern Mediterranean. These prawns are considered highly prized in Levantine fisheries, and compose most of the prawn catch off the Mediterranean coast of Egypt, being 6% of total Egyptian landings. This high abundance of invasive prawns has led to the decline of a native penaeid prawn, Melicertus kerathurus, which supported a commercial Israeli fishery throughout the 1950s. Due to outcompetition and its habitat being overrun by these migrants, this native species has since disappeared, with resultant detrimental impacts on the commercial fishery.
Recent media