Respect all members: no insults, harassment, or hate speech.
Be tolerant of different viewpoints, cultures, and beliefs. If you do not agree with others, just create separate note, article or collection.
Clearly distinguish between personal opinion and fact.
Verify facts before posting, especially when writing about history, science, or statistics.
Promotional content must be published on the “Related Services and Products” page—no more than one paragraph per service. You can also create subpages under the “Related Services and Products” page and publish longer promotional text there.
Do not post materials that infringe on copyright without permission.
Always credit sources when sharing information, quotes, or media.
Be respectful of the work of others when making changes.
Discuss major edits instead of removing others' contributions without reason.
If you notice rule-breaking, notify community about it in talks.
Do not share personal data of others without their consent.
Behavioural genetics, also referred to as behaviour genetics, is a field of scientificresearch that uses geneticmethods to investigate the nature and origins of individual differences in behaviour. While the name "behavioural genetics" connotes a focus on genetic influences, the field broadly investigates the extent to which genetic and environmental factors influence individual differences, and the development of research designs that can remove the confounding of genes and environment.
Behavioural genetics was founded as a scientific discipline by Francis Galton in the late 19th century, only to be discredited through association with eugenics movements before and during World War II. In the latter half of the 20th century, the field saw renewed prominence with research on inheritance of behaviour and mental illness in humans (typically using twin and family studies), as well as research on genetically informative model organisms through selective breeding and crosses. In the late 20th and early 21st centuries, technological advances in molecular genetics made it possible to measure and modify the genome directly. This led to major advances in model organism research (e.g., knockout mice) and in human studies (e.g., genome-wide association studies), leading to new scientific discoveries[example needed].
Findings from behavioural genetic research have broadly impacted modern understanding of the role of genetic and environmental influences on behaviour. These include evidence that nearly all researched behaviours are under a significant degree of genetic influence, and that influence tends to increase as individuals develop into adulthood. Further, most researched human behaviours are influenced by a very large number of genes and the individual effects of these genes are very small. Environmental influences also play a strong role, but they tend to make family members more different from one another, not more similar.
Farmers with wheat and cattle—Ancient Egyptian art 1,422 BCE displaying domesticated animals
Selective breeding and the domestication of animals is perhaps the earliest evidence that humans considered the idea that individual differences in behaviour could be due to natural causes.[1]Plato and Aristotle each speculated on the basis and mechanisms of inheritance of behavioural characteristics.[2] Plato, for example, argued in The Republic that selective breeding among the citizenry to encourage the development of some traits and discourage others, what today might be called eugenics, was to be encouraged in the pursuit of an ideal society.[2][3] Behavioural genetic concepts also existed during the English Renaissance, where William Shakespeare perhaps first coined the phrase "nature versus nurture" in The Tempest, where he wrote in Act IV, Scene I, that Caliban was "A devil, a born devil, on whose nature Nurture can never stick".[3][4]
Modern-day behavioural genetics began with Sir Francis Galton, a nineteenth-century intellectual and cousin of Charles Darwin.[3] Galton was a polymath who studied many subjects, including the heritability of human abilities and mental characteristics. One of Galton's investigations involved a large pedigree study of social and intellectual achievement in the Englishupper class. In 1869, 10 years after Darwin's On the Origin of Species, Galton published his results in Hereditary Genius.[5] In this work, Galton found that the rate of "eminence" was highest among close relatives of eminent individuals, and decreased as the degree of relationship to eminent individuals decreased. While Galton could not rule out the role of environmental influences on eminence, a fact which he acknowledged, the study served to initiate an important debate about the relative roles of genes and environment on behavioural characteristics. Through his work, Galton also "introduced multivariate analysis and paved the way towards modern Bayesian statistics" that are used throughout the sciences—launching what has been dubbed the "Statistical Enlightenment".[6]
Galton in his later years
The field of behavioural genetics, as founded by Galton, was ultimately undermined by another of Galton's intellectual contributions, the founding of the eugenics movement in 20th century society.[3] The primary idea behind eugenics was to use selective breeding combined with knowledge about the inheritance of behaviour to improve the human species.[3] The eugenics movement was subsequently discredited by scientific corruption and genocidal actions in Nazi Germany. Behavioural genetics was thereby discredited through its association to eugenics.[3] The field once again gained status as a distinct scientific discipline through the publication of early texts on behavioural genetics, such as Calvin S. Hall's 1951 book chapter on behavioural genetics, in which he introduced the term "psychogenetics",[7] which enjoyed some limited popularity in the 1960s and 1970s.[8][9] However, it eventually disappeared from usage in favour of "behaviour genetics".
The start of behaviour genetics as a well-identified field was marked by the publication in 1960 of the book Behavior Genetics by John L. Fuller and William Robert (Bob) Thompson.[1][10] It is widely accepted now that many if not most behaviours in animals and humans are under significant genetic influence, although the extent of genetic influence for any particular trait can differ widely.[11][12] A decade later, in February 1970, the first issue of the journal Behavior Genetics was published and in 1972 the Behavior Genetics Association was formed with Theodosius Dobzhansky elected as the association's first president. The field has since grown and diversified, touching many scientific disciplines.[3][13]
The primary goal of behavioural genetics is to investigate the nature and origins of individual differences in behaviour.[3] A wide variety of different methodological approaches are used in behavioural genetic research,[14] only a few of which are outlined below.
This section needs expansion. You can help by adding to it. (February 2020)
Investigators in animal behaviour genetics can carefully control for environmental factors and can experimentally manipulate genetic variants, allowing for a degree of causal inference that is not available in studies on human behavioural genetics.[15] In animal research selection experiments have often been employed. For example, laboratory house mice have been bred for open-field behaviour,[16]thermoregulatorynesting,[17] and voluntary wheel-running behaviour.[18] A range of methods in these designs are covered on those pages.
Behavioural geneticists using model organisms employ a range of molecular techniques to alter, insert, or delete genes. These techniques include knockouts, floxing, gene knockdown, or genome editing using methods like CRISPR-Cas9.[19] These techniques allow behavioural geneticists different levels of control in the model organism's genome, to evaluate the molecular, physiological, or behavioural outcome of genetic changes.[20] Animals commonly used as model organisms in behavioural genetics include mice,[21]zebra fish,[22]Drosophila,[23] and the nematode species C. elegans.[24]
Machine learning and A.I. developments are allowing researchers to design experiments that are able to manage the complexity and large data sets generated, allowing for increasingly complex behavioural experiments.[25]
Some research designs used in behavioural genetic research are variations on family designs (also known as pedigree designs), including twin studies and adoption studies.[14]Quantitative genetic modelling of individuals with known genetic relationships (e.g., parent-child, sibling, dizygotic and monozygotic twins) allows one to estimate to what extent genes and environment contribute to phenotypic differences among individuals.[26]
The basic intuition of the twin study is that monozygotic twins share 100% of their genome and dizygotic twins share, on average, 50% of their segregating genome. Thus, differences between the two members of a monozygotic twin pair can only be due to differences in their environment, whereas dizygotic twins will differ from one another due to genes in addition to the environment. Under this simplistic model, if dizygotic twins differ more than monozygotic twins it can only be attributable to genetic influences. An important assumption of the twin model is the equal environment assumption[27] that monozygotic twins have the same shared environmental experiences as dizygotic twins. If, for example, monozygotic twins tend to have more similar experiences than dizygotic twins—and these experiences themselves are not genetically mediated through gene-environment correlation mechanisms—then monozygotic twins will tend to be more similar to one another than dizygotic twins for reasons that have nothing to do with genes.[28] While this assumption should be kept in mind when interpreting the results of twin studies, research tends to support the equal environment assumption.[29]
Twin studies of monozygotic and dizygotic twins use a biometrical formulation to describe the influences on twin similarity and to infer heritability.[26][30]
The formulation rests on the basic observation that the variance in a phenotype is due to two sources, genes and environment. More formally, , where is the phenotype, is the effect of genes, is the effect of the environment, and is a gene by environment interaction. The term can be expanded to include additive (), dominance (), and epistatic () genetic effects. Similarly, the environmental term can be expanded to include shared environment () and non-shared environment (), which includes any measurement error. Dropping the gene by environment interaction for simplicity (typical in twin studies) and fully decomposing the and terms, we now have . Twin research then models the similarity in monozygotic twins and dizygotic twins using simplified forms of this decomposition, shown in the table.[26]
Decomposing the genetic and environmental contributions to twin similarity.[26]
Type of relationship
Full decomposition
Falconer's decomposition
Perfect similarity between siblings
Monozygotic twin correlation()
Dizygotic twin correlation ()
Where is an unknown (probably very small) quantity.
The simplified Falconer formulation can then be used to derive estimates of , , and . Rearranging and substituting the and equations one can obtain an estimate of the additive genetic variance, or heritability, , the non-shared environmental effect and, finally, the shared environmental effect .[26] The Falconer formulation is presented here to illustrate how the twin model works. Modern approaches use maximum likelihood to estimate the genetic and environmental variance components.[31]
Candidate Genes. One popular approach has been to test for association candidate genes with behavioural phenotypes, where the candidate gene is selected based on some a priori theory about biological mechanisms involved in the manifestation of a behavioural trait or phenotype.[34] In general, such studies have proven difficult to broadly replicate[35][36][37][38] and there has been concern raised that the false positive rate in this type of research is high.[34][39]
Genome-wide association studies In genome-wide association studies, researchers test the relationship of millions of genetic polymorphisms with behavioural phenotypes across the genome.[33] This approach to genetic association studies is largely atheoretical, and typically not guided by a particular biological hypothesis regarding the phenotype.[33] Genetic association findings for behavioural traits and psychiatric disorders have been found to be highly polygenic (involving many small genetic effects).[40][41][42][43][44]Study results about which and to what degree various traits, IQ and language-related skills appear to be influenced by genetics[45][46] Genetic variants identified to be associated with some trait or disease through GWAS may be used to improve disease risk predictions. However, the genetic variants identified through GWAS of common genetic variants are most likely to have a modest effect on disease risk or development of a given trait. This is different from the strong genetic contribution seen in Mendelian conditions or for some rare variants that may have a larger effect on disease.
SNP heritability and co-heritability Recently, researchers have begun to use similarity between classically unrelated people at their measured single nucleotide polymorphisms (SNPs) to estimate genetic variation or covariation that is tagged by SNPs, using mixed effects models implemented in software such as genome-wide complex trait analysis (GCTA).[47][48] To do this, researchers find the average genetic relatedness over all SNPs between all individuals in a (typically large) sample, and use Haseman–Elston regression or restricted maximum likelihood to estimate the genetic variation that is "tagged" by, or predicted by, the SNPs. The proportion of phenotypic variation that is accounted for by the genetic relatedness has been called "SNP heritability".[49] Intuitively, SNP heritability increases to the degree that phenotypic similarity is predicted by genetic similarity at measured SNPs, and is expected to be lower than the true narrow-sense heritability to the degree that measured SNPs fail to tag (typically rare) causal variants.[50] The value of this method is that it is an independent way to estimate heritability that does not require the same assumptions as those in twin and family studies, and that it gives insight into the allelic frequency spectrum of the causal variants underlying trait variation.[51]
Some behavioural genetic designs are useful not to understand genetic influences on behaviour, but to control for genetic influences to test environmentally-mediated influences on behaviour.[52] Such behavioural genetic designs may be considered a subset of natural experiments,[53]quasi-experiments that attempt to take advantage of naturally occurring situations that mimic true experiments by providing some control over an independent variable. Natural experiments can be particularly useful when experiments are infeasible, due to practical or ethical limitations.[53]
A general limitation of observational studies is that the relative influences of genes and environment are confounded. A simple demonstration of this fact is that measures of 'environmental' influence are heritable.[54] Thus, observing a correlation between an environmental risk factor and a health outcome is not necessarily evidence for environmental influence on the health outcome. Similarly, in observational studies of parent-child behavioural transmission, for example, it is impossible to know if the transmission is due to genetic or environmental influences, due to the problem of passive gene–environment correlation.[53] The simple observation that the children of parents who use drugs are more likely to use drugs as adults does not indicate why the children are more likely to use drugs when they grow up. It could be because the children are modelling their parents' behaviour. Equally plausible, it could be that the children inherited drug-use-predisposing genes from their parent, which put them at increased risk for drug use as adults regardless of their parents' behaviour. Adoption studies, which parse the relative effects of rearing environment and genetic inheritance, find a small to negligible effect of rearing environment on smoking, alcohol, and marijuana use in adopted children,[55][non-primary source needed] but a larger effect of rearing environment on harder drug use.[56][non-primary source needed]
There are many broad conclusions to be drawn from behavioural genetic research about the nature and origins of behaviour.[3][59] Three major conclusions include:[3]
all behavioural traits and disorders are influenced by genes
environmental influences tend to make members of the same family more different, rather than more similar
the influence of genes tends to increase in relative importance as individuals age.
It is clear from multiple lines of evidence that all researched behavioural traits and disorders are influenced by genes; that is, they are heritable. The single largest source of evidence comes from twin studies, where it is routinely observed that monozygotic (identical) twins are more similar to one another than are same-sex dizygotic (fraternal) twins.[11][12]
The conclusion that genetic influences are pervasive has also been observed in research designs that do not depend on the assumptions of the twin method. Adoption studies show that adoptees are routinely more similar to their biological relatives than their adoptive relatives for a wide variety of traits and disorders.[3] In the Minnesota Study of Twins Reared Apart, monozygotic twins separated shortly after birth were reunited in adulthood.[60] These adopted, reared-apart twins were as similar to one another as were twins reared together on a wide range of measures including general cognitive ability, personality, religious attitudes, and vocational interests, among others.[60] Approaches using genome-wide genotyping have allowed researchers to measure genetic relatedness between individuals and estimate heritability based on millions of genetic variants. Methods exist to test whether the extent of genetic similarity (aka, relatedness) between nominally unrelated individuals (individuals who are not close or even distant relatives) is associated with phenotypic similarity.[48] Such methods do not rely on the same assumptions as twin or adoption studies, and routinely find evidence for heritability of behavioural traits and disorders.[42][44][61]
Just as all researched human behavioural phenotypes are influenced by genes (i.e., are heritable), all such phenotypes are also influenced by the environment.[11][59] The basic fact that monozygotic twins are genetically identical but are never perfectly concordant for psychiatric disorder or perfectly correlated for behavioural traits, indicates that the environment shapes human behaviour.[59]
The nature of this environmental influence, however, is such that it tends to make individuals in the same family more different from one another, not more similar to one another.[3] That is, estimates of shared environmental effects () in human studies are small, negligible, or zero for the vast majority of behavioural traits and psychiatric disorders, whereas estimates of non-shared environmental effects () are moderate to large.[11] From twin studies is typically estimated at 0 because the correlation () between monozygotic twins is at least twice the correlation () for dizygotic twins. When using the Falconer variance decomposition () this difference between monozygotic and dizygotic twin similarity results in an estimated . The Falconer decomposition is simplistic.[26] It removes the possible influence of dominance and epistatic effects which, if present, will tend to make monozygotic twins more similar than dizygotic twins and mask the influence of shared environmental effects.[26] This is a limitation of the twin design for estimating . However, the general conclusion that shared environmental effects are negligible does not rest on twin studies alone. Adoption research also fails to find large () components; that is, adoptive parents and their adopted children tend to show much less resemblance to one another than the adopted child and his or her non-rearing biological parent.[3] In studies of adoptive families with at least one biological child and one adopted child, the sibling resemblance also tends to be nearly zero for most traits that have been studied.[11][62]
Similarity in twins and adoptees indicates a small role for shared environment in personality.
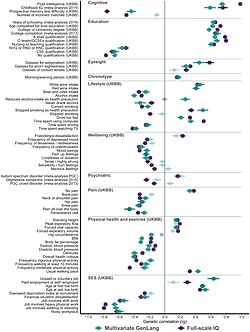
The figure provides an example from personality research, where twin and adoption studies converge on the conclusion of zero to small influences of shared environment on broad personality traits measured by the Multidimensional Personality Questionnaire including positive emotionality, negative emotionality, and constraint.[63]
Given the conclusion that all researched behavioural traits and psychiatric disorders are heritable, biological siblings will always tend to be more similar to one another than will adopted siblings. However, for some traits, especially when measured during adolescence, adopted siblings do show some significant similarity (e.g., correlations of .20) to one another. Traits that have been demonstrated to have significant shared environmental influences include internalizing and externalizing psychopathology,[64] substance use[65][non-primary source needed] and dependence,[56][non-primary source needed] and intelligence.[65][non-primary source needed]
Genetic effects on human behavioural outcomes can be described in multiple ways.[26] One way to describe the effect is in terms of how much variance in the behaviour can be accounted for by alleles in the genetic variant, otherwise known as the coefficient of determination or . An intuitive way to think about is that it describes the extent to which the genetic variant makes individuals, who harbour different alleles, different from one another on the behavioural outcome. A complementary way to describe effects of individual genetic variants is in how much change one expects on the behavioural outcome given a change in the number of risk alleles an individual harbours, often denoted by the Greek letter (denoting the slope in a regression equation), or, in the case of binary disease outcomes by the odds ratio of disease given allele status. Note the difference: describes the population-level effect of alleles within a genetic variant; or describe the effect of having a risk allele on the individual who harbours it, relative to an individual who does not harbour a risk allele.[66]
On the other hand, when assessing effects according to the metric, there are a large number of genetic variants that have very large effects on complex behavioural phenotypes. The risk alleles within such variants are exceedingly rare, such that their large behavioural effects impact only a small number of individuals. Thus, when assessed at a population level using the metric, they account for only a small amount of the differences in risk between individuals in the population. Examples include variants within APP that result in familial forms of severe early onset Alzheimer's disease but affect only relatively few individuals. Compare this to risk alleles within APOE, which pose much smaller risk compared to APP, but are far more common and therefore affect a much greater proportion of the population.[76]
Finally, there are classical behavioural disorders that are genetically simple in their etiology, such as Huntington's disease. Huntington's is caused by a single autosomaldominant variant in the HTT gene, which is the only variant that accounts for any differences among individuals in their risk for developing the disease, assuming they live long enough.[77] In the case of genetically simple and rare diseases such as Huntington's, the variant and the are simultaneously large.[66]
Behavioural genetic research and findings have at times been controversial. Some of this controversy has arisen because behavioural genetic findings can challenge societal beliefs about the nature of human behaviour and abilities. Major areas of controversy have included genetic research on topics such as racial differences, intelligence, violence, and human sexuality.[78] Other controversies have arisen due to misunderstandings of behavioural genetic research, whether by the lay public or the researchers themselves.[3] For example, the notion of heritability is easily misunderstood to imply causality, or that some behaviour or condition is determined by one's genetic endowment.[79] When behavioural genetics researchers say that a behaviour is X% heritable, that does not mean that genetics causes, determines, or fixes up to X% of the behaviour. Instead, heritability is a statement about genetic differences correlated with trait differences on the population level.[citation needed]
Historically, perhaps the most controversial subject has been on race and genetics.[78]Race is not a scientifically exact term, and its interpretation can depend on one's culture and country of origin.[80] Instead, geneticists use concepts such as ancestry, which is more rigorously defined.[81] For example, a so-called "Black" race may include all individuals of relatively recent African descent ("recent" because all humans are descended from African ancestors). However, there is more genetic diversity in Africa than the rest of the world combined,[82] so speaking of a "Black" race is without a precise genetic meaning.[81]
Qualitative research has fostered arguments that behavioural genetics is an ungovernable field without scientific norms or consensus, which fosters controversy. The argument continues that this state of affairs has led to controversies including race, intelligence, instances where variation within a single gene was found to very strongly influence a controversial phenotype (e.g., the "gay gene" controversy), and others. This argument further states that because of the persistence of controversy in behaviour genetics and the failure of disputes to be resolved, behaviour genetics does not conform to the standards of good science.[83]
The scientific assumptions on which parts of behavioural genetic research are based have also been criticized as flawed.[79] Genome wide association studies are often implemented with simplifying statistical assumptions, such as additivity, which may be statistically robust but unrealistic for some behaviours. Critics further contend that, in humans, behaviour genetics represents a misguided form of genetic reductionism based on inaccurate interpretations of statistical analyses.[84] Studies comparing monozygotic (MZ) and dizygotic (DZ) twins assume that environmental influences will be the same in both types of twins, but this assumption may also be unrealistic. MZ twins may be treated more alike than DZ twins,[79] which itself may be an example of evocative gene–environment correlation, suggesting that one's genes influence their treatment by others. It is also not possible in twin studies to eliminate effects of the shared womb environment, although studies comparing twins who experience monochorionic and dichorionic environments in utero do exist, and indicate limited impact.[85] Studies of twins separated in early life include children who were separated not at birth but part way through childhood.[79] The effect of early rearing environment can therefore be evaluated to some extent in such a study, by comparing twin similarity for those twins separated early and those separated later.[60]
^Stigler SM (July 2010). "Darwin, Galton and the Statistical Enlightenment". Journal of the Royal Statistical Society, Series A. 173 (3): 469–482. doi:10.1111/j.1467-985X.2010.00643.x. S2CID53333238.
^Hall CS (1951). "The genetics of behavior". In Stevens SS (ed.). Handbook of Experimental Psychology. New York: John Wiley and Sons. pp. 304–329.
^Grigorenko EL, Ravich-Shcherbo I (1997). "Russian psychogenetics". In Grigorenko EL (ed.). Psychology of Russia: Past, Present, Future. Commack, NY: Nova Science. pp. 83–124.
^ abPlomin R, DeFries JC, Knopik VS, Neiderhiser M (24 September 2012). Behavioral Genetics. Worth Publishers. ISBN978-1-4292-4215-8. Archived from the original on 31 December 2013. Retrieved 27 January 2016.
^Cryan JF, Holmes A (September 2005). "The ascent of mouse: advances in modelling human depression and anxiety". Nature Reviews. Drug Discovery. 4 (9): 775–790. doi:10.1038/nrd1825. PMID16138108. S2CID18207374.
^Jinks JL, Fulker DW (1970). "Comparison of the biometrical genetical, MAVA, and classical approaches to the analysis of the human behavior". Psychological Bulletin. 73 (5): 311–349. doi:10.1037/h0029135. PMID5528333.
^D'Onofrio BM, Turkheimer EN, Eaves LJ, Corey LA, Berg K, Solaas MH, et al. (November 2003). "The role of the children of twins design in elucidating causal relations between parent characteristics and child outcomes". Journal of Child Psychology and Psychiatry, and Allied Disciplines. 44 (8): 1130–44. doi:10.1111/1469-7610.00196. PMID14626455.
^Burt SA (July 2009). "Rethinking environmental contributions to child and adolescent psychopathology: a meta-analysis of shared environmental influences". Psychological Bulletin. 135 (4): 608–37. doi:10.1037/a0015702. PMID19586164.
^Corder EH, Saunders AM, Risch NJ, Strittmatter WJ, Schmechel DE, Gaskell PC, et al. (June 1994). "Protective effect of apolipoprotein E type 2 allele for late onset Alzheimer disease". Nature Genetics. 7 (2): 180–4. doi:10.1038/ng0694-180. PMID7920638. S2CID11137478.
^Luczak SE, Glatt SJ, Wall TL (July 2006). "Meta-analyses of ALDH2 and ADH1B with alcohol dependence in Asians". Psychological Bulletin. 132 (4): 607–21. doi:10.1037/0033-2909.132.4.607. PMID16822169.
Crusio WE, Gerlai RT, eds. (1999). Handbook of Molecular-Genetic Techniques for Brain and Behavior Research. Techniques in the Behavioral and Neural Sciences. Vol. 13. Elsevier. ISBN978-0-444-50239-1.
Crusio WE, Sluyter F, Gerlai RT, Pietropaolo S, eds. (2013). Behavioral Genetics of the Mouse: Genetics of Behavioral Phenotypes. Cambridge Handbooks in Behavioral Genetics. Vol. 1. Cambridge University Press. ISBN978-1-107-03481-5.
Maxson SC (10 October 2012). "Chapter 1: Behavioral Genetics". In Weiner IB, Nelson RJ, Mizumori S (eds.). Handbook of Psychology (Volume 3: Behavioral Neuroscience). John Wiley & Sons. ISBN978-0-470-89059-2. Archived from the original on 16 December 2013. Retrieved 15 December 2013.
Spinath FM, Johnson W (2011). "Chapter 10: Behavior Genetics". In Chamorro-Premuzic T, von Stumm S, Furnham A (eds.). The Wiley-Blackwell Handbook of Individual Differences. United Kingdom: Blackwell Publishing Ltd. doi:10.1002/9781444343120. ISBN978-1-4443-3438-8.
Behavioural genetics is the scientific discipline that investigates the extent to which genetic and environmental factors contribute to individual differences in human and animal behaviour, employing quantitative methods to partition variance into heritable and non-heritable components.[1][2] Pioneered by Francis Galton in the late 19th century through early inquiries into heredity and mental abilities, the field developed via twin, adoption, and family studies to estimate heritability—the proportion of phenotypic variance due to genetic variance—which consistently reveals substantial genetic influences on complex traits.[3][4]Key replicated findings include that all measured psychological traits, from cognitive abilities to personality dimensions and psychiatric disorders, show average heritability of approximately 50%, with genetic effects arising from many variants of small effect rather than single genes.[2][5] Advances in genome-wide association studies (GWAS) have identified thousands of genetic loci associated with behavioural outcomes, enabling polygenic scores that predict up to 10-15% of variance in traits like educational attainment and risk for conditions such as schizophrenia, shifting focus from broad heritability to molecular mechanisms.[4][6]The field has faced controversies, including historical associations with eugenics and ongoing debates over interpretations amid institutional biases that downplay genetic causation in favour of environmental explanations, yet empirical evidence from diverse methodologies robustly supports causal genetic roles in behavioural variation, challenging purely nurture-based models.[2][7] These insights inform applications in medicine, education, and policy by highlighting how genetic predispositions interact with environments, though shared family environments contribute minimally to variance compared to non-shared experiences and genetics.[8]
Definition and Scope
Core Principles and Terminology
Behavioral genetics examines the extent to which variation in behavioral traits within a population arises from genetic differences versus environmental influences, emphasizing quantitative methods to partition observed phenotypic variance. The core principle is that phenotypic variance, denoted as Var(P), can be decomposed into genetic and environmental components, assuming linearity and additivity in basic models: Var(P) = Var(G) + Var(E), where G represents genetic factors and E environmental factors. This decomposition rests on the foundational idea that individual differences in behavior, such as intelligence or personality, are polygenic—influenced by many genes of small effect—and substantially heritable, with estimates typically ranging from 30% to 50% across traits.[2][9]Key terminology includes phenotype, the observable behavioral trait or measure, and genotype, the underlying genetic configuration contributing to it. Heritability quantifies the proportion of phenotypic variance attributable to genetic variance: narrow-sense heritability (h²) focuses on additive genetic variance (Var(A)), while broad-sense heritability (H²) encompasses all genetic effects including dominance and epistasis. In the standard ACE model, variance is further partitioned as Var(P) = a² + c² + e², where a² is additive genetic variance, c² is shared environmental variance (common to family members), and e² is non-shared environmental variance (unique to individuals, including measurement error). This model, derived from twin and family studies, reveals that shared environment often explains little variance in behavioral traits after adolescence, with non-shared effects dominating alongside genetics.[2][9]Additional principles highlight that no behavioral traits are fully heritable or environmentally determined; genetic influences amplify over development for many traits, and gene-environment interactions (GxE) and correlations (rGE) complicate simple partitions, though basic estimates assume their minimal role for identifiability. Polygenicity implies that single genes rarely account for substantial variance, favoring genome-wide association studies (GWAS) for identification. Heritability estimates are population-specific, varying with environmental homogeneity—higher in stable environments where genetic differences more readily manifest—and do not imply determinism or immutability, as they describe variance, not individual causation.[2][5]
Distinction from Eugenics and Related Fields
Behavioural genetics is a scientific field focused on partitioning variance in behavioural traits into genetic and environmental components through empirical methods such as twin and adoption studies, without advocating for human intervention in reproduction.[7] In contrast, eugenics, coined by Francis Galton in 1883, promoted deliberate efforts to enhance human genetic quality via selective breeding, including positive incentives for reproduction among deemed "fit" individuals and negative measures like sterilization of the "unfit," which led to over 60,000 forced sterilizations in the United States between 1907 and the 1970s.[10] This prescriptive ideology, rooted in early hereditarian views, diverged from the descriptive aims of behavioural genetics by prioritizing societal engineering over neutral inquiry into causal mechanisms.[11]Although both fields trace origins to Galton's pioneering work in biometrics and heritability estimation in the late 19th century, modern behavioural genetics rejects eugenics' coercive applications, particularly after their ethical discrediting post-World War II due to associations with Nazi programs that sterilized or euthanized hundreds of thousands.[7] Professional bodies like the Behavior Genetics Association affirm that research findings should not justify discriminatory policies, emphasizing instead applications in personalized education, mental health interventions, and voluntary reproductive choices informed by polygenic risk scores.[12] Critics sometimes conflate the two due to shared interest in genetic influences on traits like intelligence, but behavioural genetics maintains a commitment to falsifiable hypotheses and environmental moderation, avoiding eugenics' deterministic and hierarchical prescriptions.[13]Distinctions from related fields include quantitative genetics, which provides statistical frameworks like Falconer's formula for heritability (h² = 2(r_MZ - r_DZ)) but applies broadly beyond behavior, and psychiatric genetics, which targets disorder-specific loci via genome-wide association studies (GWAS) rather than population-level selection.[7] Unlike eugenics' focus on aggregate population improvement, these fields prioritize mechanistic understanding, such as gene-environment interactions, to explain variance without implying inherent superiority or policy mandates for breeding.[11] Empirical data from large-scale GWAS, identifying thousands of variants for traits like educational attainment with effect sizes around 0.1% per SNP, underscore the polygenic, probabilistic nature of inheritance, incompatible with eugenics' simplified Mendelian assumptions prevalent in early 20th-century implementations.[11]
Historical Development
Early Pioneers and Quantitative Foundations (19th-early 20th Century)
Francis Galton, inspired by Charles Darwin's On the Origin of Species published in 1859, initiated systematic inquiry into the inheritance of human mental and behavioral traits in the 1860s.[3] In his 1869 book Hereditary Genius, Galton analyzed the pedigrees of eminent individuals, concluding that intellectual ability is substantially heritable, with parental eminence predicting offspring success at rates far exceeding chance.[14] He employed early statistical methods, including deviation measures from population averages, to quantify resemblance among relatives, laying groundwork for partitioning phenotypic variance into genetic and environmental components.[15]Galton's 1875 publication "The History of Twins" marked the inception of twin-based research in behavioral genetics, using case studies of reared-apart and reared-together twins to disentangle hereditary from environmental influences on traits like temperament and ability.[16][17] Although he did not explicitly contrast monozygotic and dizygotic twins as later formalized, Galton observed that similarities in identical twins persisted despite divergent upbringings, attributing this to innate factors over shared nurture.[18] His 1889 work Natural Inheritance advanced quantitative models, introducing the concept of regression toward the mean in hereditary transmission and proposing the law of ancestral heredity, which weighted contributions from ancestors proportionally to their generational proximity (e.g., parents 1/2, grandparents 1/4).[19]Karl Pearson, Galton's protégé and collaborator from the 1890s, formalized these ideas through biometric methods, developing the product-moment correlation coefficient in 1895 to measure trait covariation across relatives.[20] Pearson's series of papers, beginning with "Mathematical Contributions to the Theory of Evolution" in 1893, refined regression and heredity models, applying them to continuous traits like height and intelligence, and defending gradualist, polygenic inheritance against emerging Mendelian particulate views.[21] By the early 1900s, Pearson established the Biometrika journal in 1901 and the Galton Laboratory, institutionalizing quantitative genetics and enabling estimation of heritability as twice the parent-offspring correlation for such traits.[22] These foundations emphasized empirical measurement of familial resemblances to infer genetic variance, influencing subsequent debates on the polygenic basis of behavioral differences.[23]
Twin Studies and Mid-20th Century Revival (1920s-1970s)
Twin studies emerged in the 1920s as a primary method in behavioral genetics to partition variance in traits like intelligence into genetic and environmental components by comparing resemblances between monozygotic (MZ) twins, who share nearly 100% of their genes, and dizygotic (DZ) twins, who share about 50% on average.[24] Curtis Merriman's 1924 study represented an early application, analyzing mental test scores from 33 twin pairs and finding significantly lower intra-pair variance in MZ twins compared to DZ twins, implying a genetic contribution to intellectual resemblance.[24] This approach built on quantitative genetic principles, using the difference in twin correlations to estimate broad-sense heritability, with Merriman's data suggesting heritability around 0.6 for mental ability.[25]Subsequent work in the late 1920s expanded sample sizes and traits examined. Freeman, Holzinger, and Mitchell's 1928 investigation of over 400 twin pairs assessed intelligence, school achievement, and conduct, reporting MZ correlations for IQ exceeding 0.85 while DZ correlations were around 0.60, yielding heritability estimates of approximately 0.50 after accounting for shared environment.[26] These studies faced limitations, including small samples, imprecise zygosity determination via physical resemblance rather than genetic markers, and potential selective ascertainment, yet they established the twin method's utility in demonstrating that environmental differences alone could not explain observed similarities in MZ pairs.[4]The 1930s saw innovations like studies of twins reared apart to further isolate genetic effects. Newman, Freeman, and Holzinger's 1937 analysis of 19 MZ twin pairs separated early in life found an IQ correlation of 0.68 despite diverse rearing environments, with mean absolute IQ differences of only 9.2 points, lower than typical sibling differences and supporting heritability estimates over 0.70 when combined with reared-together data. Applications extended to psychopathology; Franz Kallmann's twin studies in the 1940s, culminating in his 1946 report on 691 schizophrenic index cases, documented MZ concordance rates of 85.8% for chronic schizophrenia versus 14.3% for DZ twins, indicating strong familial aggregation consistent with polygenic inheritance rather than simple Mendelian patterns.[27] Kallmann's work, though later critiqued for diagnostic inconsistencies and overestimation of concordance due to broad phenotyping, provided early empirical evidence against purely environmental models of mental disorders.[28]Post-World War II, behavioral genetics encountered skepticism amid associations with eugenics and revelations of unethical twin experiments under the Nazis, leading to a temporary decline in human studies and dominance of environmentalist paradigms like behaviorism.[29] Nonetheless, quantitative foundations persisted, with revival accelerating in the 1950s and 1960s through longitudinal designs and refined statistical models. The Louisville Twin Study, initiated in 1957 with over 500 pairs, tracked cognitive development from infancy, revealing heritability of mental speed and IQ rising from near zero in early childhood to 0.80 by adolescence, as shared environmental effects diminished and genetic variances amplified with age.[30] This pattern underscored developmental gene-environment interplay, where initial environmental dominance gave way to genetic expression as individuals selected environments aligning with genotypes.[31]By the 1970s, accumulating twin data across traits—yielding heritability estimates of 0.40-0.50 for personality dimensions like extraversion and neuroticism—challenged strict nurture doctrines and spurred institutional growth, including dedicated training programs.[4] Methodological improvements, such as serological zygosity confirmation and model-fitting via maximum likelihood estimation, enhanced precision, distinguishing additive genetic, dominance, and shared environmental variances.[32] These advances, grounded in empirical correlations rather than ideology, substantiated causal genetic roles in behavioral variation while acknowledging non-shared environmental influences on individual differences.[3]
Molecular Revolution and GWAS Era (1980s-Present)
The molecular revolution in behavioral genetics commenced in the 1980s, driven by advances in recombinant DNA technology, DNA sequencing, and the ability to clone and manipulate genes, which shifted research from quantitative estimates of heritability to identifying specific genetic variants associated with behavioral traits.[4] These tools, including restriction fragment length polymorphisms (RFLPs) as early markers, enabled initial linkage studies in families for monogenic disorders with behavioral components, such as Huntington's disease, but proved challenging for polygenic behavioral phenotypes due to their complex, non-Mendelian inheritance.[33] By the late 1980s and into the 1990s, the field embraced candidate gene association studies, hypothesizing that variants in genes related to neurotransmitter systems (e.g., dopamine receptor D4 for novelty-seeking or monoamine oxidase A for aggression) influenced traits like personality and impulsivity; however, thousands of such studies yielded low replication rates, often attributable to small sample sizes, inadequate statistical power, population stratification, and overemphasis on functional plausibility without genome-wide context.[4][5]The genome-wide association study (GWAS) era, enabled by high-density single nucleotide polymorphism (SNP) arrays around 2005, marked a paradigm shift toward hypothesis-free, whole-genome scans requiring large consortia to detect small-effect common variants.[4] Initial applications to psychiatric disorders yielded the first replicated loci in 2007–2008, such as for bipolar disorder and schizophrenia, where the Psychiatric GWAS Consortium identified common alleles at loci like MHC and CACNA1C, explaining modest fractions of liability (e.g., ~3% for schizophrenia by 2009).[34][35] For non-clinical behavioral traits, GWAS progressed more slowly due to measurement challenges and lower heritability signals, but by the 2010s, studies on educational attainment (a proxy for cognitive ability) in samples exceeding 100,000 identified hundreds of loci, with polygenic scores (PRS) aggregating effects to predict 10–15% of variance in independent samples.[4] Similarly, GWAS for personality traits like neuroticism revealed polygenic architectures overlapping with psychopathology, with recent meta-analyses (e.g., 2024 for Big Five traits in ~224,000 individuals) confirming small but robust effects across thousands of SNPs.[36]GWAS findings validated twin-study heritability through SNP-based estimates (via methods like GREML), resolving much of the "missing heritability" paradox by demonstrating that behavioral traits arise from myriad common variants of tiny effect rather than rare high-impact mutations, though rare variants and non-additive interactions remain underexplored.[37] Polygenic risk scores, first applied effectively to schizophrenia in 2009 and extended to normal-range behaviors by the mid-2010s, enable within-population prediction and causal inference via Mendelian randomization, revealing genetic correlations (e.g., between intelligence and mental health) that underscore pleiotropy.[6][4] Despite successes, challenges persist: PRS explain only a subset of twin heritability (e.g., <20% for most traits), exhibit portability issues across ancestries due to linkage disequilibrium differences, and face ascertainment biases in samples favoring European-descent populations, prompting calls for diverse genotyping.[38] Ongoing mega-consortia, such as those under the PGC and SSGAC, continue scaling to millions of genomes, integrating functional genomics to link variants to brain expression and causal pathways.[34]
Methodological Approaches
Quantitative Genetic Designs
Quantitative genetic designs in behavioral genetics utilize statistical analyses of phenotypic similarities among relatives with differing genetic relatedness to estimate the proportions of variance attributable to genetic factors, shared environments, and nonshared environments, without requiring identification of specific genetic variants. These approaches, rooted in quantitative genetics principles developed in the early 20th century, model continuous traits as the sum of additive genetic effects (A), dominance or epistatic effects (D), shared environmental effects (C), and unique environmental effects (E), where phenotypic variance Var(P) = A + D + C + E, and heritability in the broad sense is (A + D)/Var(P).[4] Such designs provide population-level estimates of genetic influence, assuming random mating, no genotype-environment interaction or correlation unless modeled, and, crucially for twin-based methods, environmental similarity between monozygotic (MZ) and dizygotic (DZ) twins comparable to their genetic resemblance. Empirical tests, including direct assessments of MZ-DZ environmental differences, support this equal environments assumption for most behavioral traits, countering critiques that overestimate environmental confounding.[2]Twin studies form the cornerstone of these designs, comparing intraclass correlations for MZ twins, who share approximately 100% of segregating genetic variants, against DZ twins, who share 50% on average, reared together. Under the ACE model—a structural equation framework fitting observed correlations to latent variance components—the MZ correlation approximates A + C, while the DZ correlation approximates A/2 + C (assuming no dominance); unique environmental effects E (including measurement error) account for within-pair differences, with total variance normalized to 1.0 such that A + C + E = 1.0.[39] Model-fitting via maximum likelihood allows testing submodels (e.g., AE by dropping C if nonsignificant) and yields confidence intervals; for instance, meta-analyses across thousands of twin pairs report median heritabilities around 0.50 for psychological traits.[2] A simpler method-of-moments estimator, Falconer's formula, approximates narrow-sense heritability as h² = 2(r_MZ - r_DZ), effective for additive models but less precise when dominance or shared environment operates.[40]Adoption and family studies complement twins by disentangling genetic from rearing effects. In adoption designs, correlations between biological relatives reared apart isolate genetic variance (approximating A), while adoptive relative correlations capture shared environment C; combined twin-adoption analyses, as in the Colorado Adoption Project initiated in 1975, refine estimates by modeling both.[4] Family studies, examining parent-offspring (r ≈ 0.5 genetic) or full sibling (r = 0.5) resemblances, provide baseline data but confound genetics with cultural transmission and shared environment, necessitating extensions like half-sibling or stepfamily comparisons for decomposition.[41] These designs collectively enable robust heritability estimates, with longitudinal extensions tracking developmental changes, such as rising genetic influence on intelligence from 20% in infancy to 80% in adulthood.[42]Advanced quantitative techniques incorporate extended kinship or genomic relatedness matrices (GRMs) from pedigree data, bridging to molecular methods, though classical designs remain foundational due to their power in large samples like the Minnesota Twin Registry (ongoing since 1983) or UK Twins Early Development Study (1994–present). Limitations include population specificity—heritabilities may vary by socioeconomic context—and inability to pinpoint causal mechanisms, addressed by integrating with quasi-experimental controls for assortative mating or gene-environment interplay.[2] Despite academic biases favoring environmental explanations, replicated findings across designs affirm substantial genetic variance for behavioral traits, informing causal realism over nurture-only narratives.[4]
Twin, Adoption, and Family Studies
Twin studies represent a primary method in quantitative behavioral genetics for estimating the relative contributions of genetic and environmental factors to phenotypic variance. In the classical twin design, monozygotic (MZ) twins, who share approximately 100% of their segregating genetic variants, are compared to dizygotic (DZ) twins, who share about 50% on average, with both types reared in the same family. The phenotypic correlation for a trait is expected to be higher in MZ pairs than DZ pairs if genetic factors are influential, allowing estimation of broad-sense heritability as twice the difference in correlations: hB2=2(rMZ−rDZ).[43] This approach assumes the equal environments assumption (EEA), positing that MZ and DZ twins experience equivalent environmental similarity relevant to the trait, and no substantial gene-environment correlation or assortative mating effects beyond additive genetics.[44] Empirical validations, including self-report measures of environmental similarity and extended family designs, indicate the EEA holds for traits like intelligence and personality, though potential violations for highly socially influenced traits can inflate heritability estimates if MZ twins select more similar environments.[45]Structural equation modeling of twin data typically decomposes variance into additive genetic (A), shared environmental (C), and unique environmental (E) components via the ACE model, where rMZ=A+C and rDZ=0.5A+C.[46] Large-scale meta-analyses aggregating over 14 million twin pairs across 2,748 studies report median heritability of 0.49 for human traits overall, with behavioral phenotypes such as cognitive abilities (around 0.50-0.80), personality dimensions (0.30-0.50), and psychiatric disorders (0.30-0.60) showing substantial genetic influence, while shared environment often explains less than 20% of variance in adulthood.[47] For instance, a meta-analysis of educational achievement from up to 5,330 MZ and 7,084 DZ pairs yielded heritability estimates of 0.66-0.73 for reading and mathematics skills.[48] These findings have been replicated across diverse populations and traits, with twin estimates aligning with molecular genetic methods like genome-wide association studies, countering claims of systematic overestimation due to violated assumptions.[2]Adoption studies disentangle genetic from shared environmental effects by comparing adoptees to their biological and adoptive relatives, where biological kin correlations reflect primarily genetic transmission, and adoptive kin correlations capture nongenetic familial influences.[49] Pioneering efforts, such as the Minnesota Study of Twins Reared Apart (initiated in 1979 with over 100 MZ pairs separated early in life), found MZ twin correlations for IQ (0.69-0.78) and personality traits comparable to those reared together, indicating minimal shared environment impact and heritability exceeding 0.70 for intelligence.[2] The Colorado Adoption Project (1975-1994, involving 245 adoptive families) similarly showed adoptee IQ correlations with biological parents (0.32 at age 12) surpassing those with adoptive parents (0.13), supporting genetic continuity while highlighting unique environmental variance.[50] Across replicated adoption designs, genetic factors consistently account for 40-60% of variance in behavioral outcomes like aggression, alcoholism risk, and cognitive performance, with adoptees' traits predicting adoptive parenting responses via evocative gene-environment correlations.[51]Family studies evaluate trait covariation among biological relatives of varying degrees of relatedness—such as parent-offspring (r ≈ 0.5 expected under full heritability) or siblings (r ≈ 0.5)—to detect aggregation, but they confound genetic and shared environmental effects, limiting precise partitioning without extensions like half-sibling comparisons. In behavioral genetics, these designs provide foundational evidence of heritability for traits like schizophrenia (familial risk ratios up to 10-fold for first-degree relatives) and inform prior probabilities for linkage analysis, though they underestimate broad heritability compared to twin methods due to unmodeled dominance or epistasis.[52] Modern family-based quantitative approaches, integrating pedigrees with genomic data, refine estimates by modeling relatedness via identity-by-descent, revealing genetic influences on social-economic outcomes like class attainment (heritability ~0.20-0.40).[53]Collectively, twin, adoption, and family studies demonstrate pervasive genetic variance in behavioral traits, with heritability typically ranging 0.30-0.80, diminishing shared environment effects post-infancy, and emphasizing nonshared experiences as key to individual differences; these quasi-experimental designs underpin causal inferences robust to many environmental confounds.[54]
Heritability Estimation Techniques
Heritability in behavioral genetics is estimated primarily through quantitative genetic designs that partition phenotypic variance into genetic and environmental components, with twin studies providing the foundational approach.[55] The classical twin method compares monozygotic (MZ) twins, who share nearly 100% of their genetic material, with dizygotic (DZ) twins, who share about 50% on average, under the assumption of equal environmental similarity for both types.[56] This design yields heritability estimates by leveraging differences in twin correlations for a trait.A straightforward technique is Falconer's formula, which calculates narrow-sense heritability as h2=2(rMZ−rDZ), where rMZ and rDZ are the intraclass correlations for MZ and DZ twins, respectively.[57] This formula assumes additive genetic effects predominate, with no significant dominance, epistasis, or gene-environment interactions, and equates shared environmental influences for MZ and DZ pairs.[58] It isolates additive genetic variance (VA) as the proportion of total phenotypic variance (VP) attributable to transmissible genetic effects, useful for predicting response to selection but potentially underestimating broad-sense heritability if non-additive effects are present.[55]More sophisticated estimation employs structural equation modeling (SEM) to fit biometric models like the ACE framework to twin covariances.[59] In the ACE model, additive genetic influences (A) explain twice the difference in MZ and DZ correlations, shared environmental effects (C) capture common family influences via C=2rDZ−rMZ, and unique environmental effects (E) account for the remainder as E=1−rMZ.[60] Heritability is then h2=A/VP, often yielding estimates of 40-60% for complex behavioral traits across meta-analyses.[61] Variants like the ADE model incorporate dominance (D) when MZ correlations substantially exceed twice DZ correlations, partitioning rMZ=A+D and rDZ=0.5A+0.25D.[62]Adoption and family studies complement twin designs by estimating heritability through comparisons of biological versus adoptive relatives, reducing shared environment confounds.[56] Parent-offspring regressions yield narrow-sense heritability as twice the regression slope, assuming no assortative mating or cultural transmission biases.[63] Sibling correlations in adoptive designs approximate half the narrow-sense heritability, with higher correlations among biological kin indicating genetic influence.[55] These methods, while less powered than twins for high-resolution partitioning, validate twin estimates and address assumptions like MZ-DZ environmental equivalence, which some critiques argue may inflate heritability if MZ twins experience more similar rearing.[64] Model selection via likelihood ratio tests or information criteria determines the best-fitting model (e.g., ACE vs. AE), ensuring estimates reflect data without overparameterization.[40] Despite limitations such as population-specific assumptions and potential violations from assortative mating, these techniques provide robust, replicable heritability figures when triangulated across designs.[48]
Molecular and Genomic Methods
Molecular and genomic methods represent a paradigm shift in behavioral genetics, enabling the identification of specific DNA variants associated with behavioral traits through direct examination of genetic sequences rather than indirect inferences from relatedness. These approaches, which gained traction in the late 20th century with advances in sequencing technology, include linkage analysis, candidate gene association studies, and genome-wide association studies (GWAS). Linkage studies initially mapped chromosomal regions co-inherited with traits in families, but their power for complex polygenic behaviors proved limited due to the diffuse nature of genetic influences.[65] By the 2000s, the field pivoted toward association methods, leveraging population samples to detect statistical links between single nucleotide polymorphisms (SNPs) and phenotypes, informed by the polygenic architecture revealed by quantitative genetics—where thousands of variants each contribute small effects to trait variance.[66] This integration of molecular data with heritability estimates from twin studies has illuminated causal pathways, though challenges persist in accounting for gene-environment interactions and the "missing heritability" gap, where identified variants explain only a fraction of twin-study estimates.[5]Candidate gene studies, prominent from the 1990s to 2010s, focused on predefined genes hypothesized to influence behaviors based on neurobiological theories, such as serotonin transporter variants (e.g., 5-HTTLPR) for anxiety or aggression. These targeted polymorphisms in genes like DRD4 for novelty-seeking or MAOA for impulsivity, often using small cohorts (typically under 1,000 participants). However, systematic reviews have documented replication rates below 10% for such associations, attributed to underpowered designs, population stratification artifacts, and selective reporting, rendering most findings non-robust upon meta-analysis.[67][68] For instance, early claims linking COMT Val158Met to executive function in schizophrenia spectra failed large-scale validation, highlighting the pitfalls of hypothesis-driven selection in polygenic systems where no single gene dominates.[69] Despite these limitations, select associations, such as CACNA1C with bipolar disorder-related traits, have withstood scrutiny when corroborated by functional studies, underscoring the method's utility for biologically plausible pathways when sample sizes exceed 10,000.[70]Genome-wide association studies, enabled by SNP arrays post-Human Genome Project (completed 2003), scan millions of markers across the genome in unrelated individuals, bypassing prior hypotheses to detect common variants with allele frequencies above 1%. Requiring massive samples—often hundreds of thousands—to overcome multiple-testing corrections (e.g., Bonferroni threshold of 5×10^{-8}), GWAS have robustly identified loci for behavioral traits; for example, a 2017 meta-analysis of over 60,000 participants pinpointed six novel SNPs for neuroticism, collectively explaining ~1-2% of variance but aligning with twin heritability of ~40%.[71] Recent consortia, like those from the Social Science Genetic Association Consortium, have extended this to educational attainment and cognitive ability, with 2022 analyses of millions yielding thousands of hits that, via polygenic risk scores (PRS), predict ~10-15% of population variance in intelligence.[72] PRS aggregate weighted effects from GWAS hits into a single metric, demonstrating cross-trait pleiotropy—e.g., schizophrenia PRS correlating with creativity measures—and longitudinal utility, such as forecasting adolescent antisocial behavior beyond socioeconomic factors.[73] Nonetheless, GWAS efficacy is tempered by European-ancestry bias in training data, reducing transferability to diverse populations (e.g., PRS accuracy drops 50% in African cohorts), and the predominance of common variants, leaving rare and structural variants underexplored.[74] Emerging whole-genome sequencing addresses this, promising fuller capture of heritability as sample sizes scale to millions.[38]
Candidate Gene Studies and Early Associations
Candidate gene studies in behavioral genetics involve hypothesis-driven investigations targeting specific genes selected based on their presumed biological relevance to behavioral traits, such as those encoding proteins in neurotransmitter systems like dopamine, serotonin, or monoamine oxidase.[75] These approaches gained prominence in the 1990s following advances in genotyping techniques and the identification of functional polymorphisms, allowing researchers to test associations between variants in predefined "candidate" genes and phenotypes like personality traits, cognitive abilities, or psychiatric disorders.[5] Early efforts focused on small-scale association studies, often with sample sizes under 500 participants, examining main effects or gene-environment interactions (G×E).[67]Pioneering associations included the 1996 report linking the 7-repeat allele of the dopamine D4 receptor gene (DRD4) to novelty-seeking behavior in a sample of 315 individuals, a finding initially replicated in some follow-ups but later contested due to inconsistent effect sizes across populations.[76] Similarly, the 2002 study by Caspi et al. identified a G×E interaction between the monoamine oxidase A (MAOA) low-activity variant and childhood maltreatment predicting antisocial behavior in a New Zealand cohort of over 1,000 males, which spurred interest in gene-environment interplay but faced replication challenges in larger datasets.[67] Other early candidates targeted serotonin transporter gene (5-HTTLPR) polymorphisms with anxiety-related traits and the catechol-O-methyltransferase (COMT Val158Met) variant with executive function and prefrontal cortex activity, drawing from animal models and pharmacological evidence.[77] By the early 2000s, over 1,000 such studies had been published in psychiatry and psychology, with dopamine and serotonin pathway genes comprising the majority.[5]Despite initial enthusiasm, candidate gene studies suffered from systemic limitations, including underpowered designs prone to false positives, inadequate correction for multiple testing, and publication bias favoring significant results.[78] Meta-analyses and large-scale replication attempts, such as those aggregating over 100 G×E findings from 2000–2009, revealed that fewer than 5% held up robustly, with many "hits" vanishing in independent samples exceeding 10,000 participants.[67] For instance, a 2019 analysis of depression candidate genes across multiple cohorts totaling over 40,000 individuals found no support for historical associations like 5-HTT or BDNF variants, attributing prior reports to inflated effect sizes from small studies (odds ratios often <1.2 in true positives).[68] These failures underscored the polygenic nature of behavioral traits, where individual variants explain negligible variance (<0.1% on average), prompting a pivot to agnostic genome-wide association studies (GWAS) by the mid-2000s.[77] Reviews emphasize that while candidate approaches provided mechanistic hypotheses, their empirical yield was minimal, highlighting the need for stringent preregistration and larger consortia to mitigate bias.[75]
Genome-Wide Association Studies (GWAS) and Polygenic Risk Scores
Genome-wide association studies (GWAS) systematically scan the genomes of large cohorts to identify single nucleotide polymorphisms (SNPs) associated with complex traits or disorders by testing millions of variants for statistical associations with phenotypic outcomes.[79] In behavioral genetics, GWAS have been pivotal since the mid-2000s, with initial applications to psychiatric conditions like schizophrenia emerging around 2007, followed by larger consortia efforts that identified genome-wide significant loci through meta-analyses of tens of thousands of cases.[80] These studies reveal that behavioral traits, including intelligence and psychopathology, exhibit polygenic architectures involving thousands of common variants each contributing small effects, often explaining only a fraction of twin-study heritability due to factors like rare variants and gene-environment interactions not captured in initial scans.[81] For instance, schizophrenia GWAS have pinpointed over 100 loci, with sample sizes exceeding 56,000 cases and 78,000 controls by 2021, demonstrating genetic overlap with bipolar disorder and negative correlations with intelligence (genetic correlation rg=−0.14).[82][83]Polygenic risk scores (PRS), derived from GWAS summary statistics, aggregate weighted effects of trait-associated SNPs to estimate an individual's genetic liability for a behavioral outcome.[84] In practice, PRS are constructed by pruning and thresholding SNPs for linkage disequilibrium, then summing allele dosages multiplied by effect sizes from discovery samples, enabling prediction in independent cohorts. For behavioral traits, PRS have shown modest but replicable predictive power; for example, schizophrenia PRS explain 7-10% of variance in case-control status and correlate with subclinical symptoms in the general population.[85] Intelligence PRS, based on GWAS of educational attainment proxies, account for up to 10-15% of phenotypic variance in European-ancestry samples as of recent large-scale studies, with applications extending to longitudinal outcomes like academic achievement.[81] Similarly, PRS for traits like neuroticism or antisocial behavior predict personality dimensions and adolescent conduct issues beyond environmental factors, highlighting causal genetic influences on development.[73][86]Despite advances, GWAS and PRS face challenges including ascertainment biases in psychiatric samples, which may inflate overlap estimates, and limited transferability across ancestries due to linkage disequilibrium differences, with most discovery data from European populations.[87] SNP-based heritability from GWAS (e.g., 20-30% for schizophrenia) remains below twin estimates (around 80%), attributed partly to uncaptured rare variants and non-additive effects, though increasing sample sizes continue to narrow this gap.[88] These tools underscore the polygenic, omnigenic nature of behavioral genetics, where pervasive weak effects across the genome contribute to trait variation, informing causal models over candidate gene approaches that yielded few replicable hits.[89]
Animal and Experimental Models
Animal models in behavioral genetics enable precise manipulation of genetic factors under controlled environments, providing causal evidence for heritability that complements observational human studies. Selective breeding experiments, initiated as early as the 19th century but refined in the 20th, demonstrate high heritability for behavioral traits by divergently selecting populations for extremes of a phenotype, such as voluntary wheel-running in mice, where high-activity lines exhibit correlated increases in exploratory behavior and reduced anxiety-like responses after over 60 generations of selection.[90] In rats, bidirectional selection for emotional reactivity has produced strains differing markedly in open-field exploration and avoidance learning, with heritability estimates exceeding 0.30 for these traits.[91]Inbred strains of rodents, particularly mice, form the backbone of experimental designs by minimizing genetic variation within strains while maximizing differences between them; for instance, the C57BL/6 strain shows robust learning in maze tasks compared to DBA/2, underscoring genetic contributions to cognitive behaviors.[92] These strains facilitate quantitative genetic analyses, revealing that most behavioral variations, from aggression to sociability, have substantial genetic components, often with narrow-sense heritabilities around 0.20-0.50 across traits.[29]Molecular approaches, including gene knockouts and transgenics, allow targeted disruption or overexpression to isolate gene effects on behavior; knockout of the serotonin transporter gene (SERT) in mice yields anxiety- and depression-like phenotypes, including increased immobility in forced-swim tests, depending on genetic background strains like 129S6 or C57BL/6.[93] Similarly, inactivation of genes like MAOA or NOS1 in mice elevates aggression, as measured by resident-intruder assays, supporting causal roles in impulsive behaviors.[94]Recent advances with CRISPR-Cas9 enable multiplexed editing in model organisms like mice and zebrafish, producing precise mutations to model polygenic traits; for example, editing vasopressin receptor genes in rodents alters social recognition and affiliation, with edited mice showing reduced partner preference in three-chamber tests.[95] In Drosophila, CRISPR knockouts of foraging gene variants shift feeding behaviors, illustrating gene-environment interactions in simple neural circuits.[92] These models, while powerful for mechanistic insights—such as circuit-level changes in anxiety via optogenetics in knockouts—face translational challenges due to species-specific neural architectures, though convergent findings across models bolster confidence in genetic causality for human analogs.[96][97]
Quasi-Experimental and Causal Inference Methods
Quasi-experimental designs in behavioral genetics leverage natural genetic and familial structures to approximate randomized experiments, enabling stronger causal inferences about genetic and environmental influences on behavior without ethical violations inherent in manipulating human traits. These methods address confounding by exploiting quasi-random assignment, such as monozygotic twins' identical genetics or siblings' shared rearing environments, to isolate causal pathways from associations. For instance, sibling comparison designs use fixed-effects models within families to control for unobserved confounders like parental genetics and socioeconomic status, revealing that genetic differences between siblings predict behavioral outcomes beyond shared environments.[98] Such approaches have demonstrated, for example, reduced bias in estimating direct genetic effects on educational attainment when comparing within-sibship genome-wide association results to population-level estimates.[99]Family-based quasi-experimental methods, including adoption and extended kinship designs, further disentangle causal transmission across generations by comparing offspring outcomes relative to biological versus adoptive parents. In adoption studies, children raised apart from biological relatives help parse genetic from rearing effects; analyses show that genetic propensities for externalizing behaviors persist despite adoptive environments, supporting causal genetic roles.[100] Extended family designs, such as children-of-twins or cousin comparisons, extend this by incorporating varying degrees of relatedness to test intergenerational causality, often finding that genetic correlations between parental traits and offspring behavior exceed environmental ones.[101] These designs mitigate population stratification biases and have been applied to psychopathology, where they confirm genetic causation in substance use disorders by showing within-family associations persist after controlling for familial confounds.[102]Mendelian randomization (MR) represents a genomic quasi-experimental technique using germline genetic variants as instrumental variables to infer causality between modifiable exposures and behavioral outcomes, assuming variants are randomly assorted at conception and free from pleiotropy or population confounding. In behavioral genetics, MR has tested causal links, such as body mass index's effect on cognitive function, finding bidirectional influences where higher BMI genetically predicts lower cognition, consistent with observational data but robust to reverse causation.[103] Applications in psychiatry reveal, for example, that genetically predicted neuroticism causally increases depression risk, with odds ratios around 1.2-1.5 per standard deviation, while ruling out causality for some hypothesized environmental mediators.[104] Sensitivity analyses, like MR-Egger for pleiotropy, strengthen inferences; violations, such as horizontal pleiotropy, are assessed via heterogeneity tests, ensuring reliability in polygenic contexts.[105]Multivariate extensions of these methods integrate genetic data with longitudinal or co-twin controls to test quasi-causal hypotheses, such as gene-environment interplay. For instance, within-twin pair analyses of polygenic scores have isolated causal genetic effects on personality traits, showing stability over development independent of shared environments.[106] Challenges include weak instrument bias from polygenic small effects and assumptions of no assortative mating, addressed via two-sample MR or family-stratified designs; despite limitations, these approaches have advanced causal knowledge by falsifying non-genetic explanations for heritable behavioral variances.[107]
Key Empirical Findings
Pervasiveness of Genetic Influences Across Traits
Twin and family studies have established that genetic factors contribute substantially to individual differences across a broad spectrum of human traits, including behavioral phenotypes. A comprehensive meta-analysis of 17,804 traits, drawn from 2,748 publications involving 14,558,903 twin pairs, yielded a median broad-sense heritability estimate of 49%, with significant genetic influences for 69% of traits.[47] These estimates cluster by functional domains, such as anthropometrics, cardiovascular, and psychiatric categories, reflecting systematic patterns of genetic and environmental variance components rather than random variation.[47] For most traits, models consistent with additive genetic effects predominate, with limited evidence for substantial non-additive genetics or shared environment across domains.[47]Within behavioral genetics, this pervasiveness manifests as ubiquitous genetic influences on psychological traits, a finding replicated across twin, adoption, and molecular designs worldwide. No adequately powered behavioral genetic study has reported heritability near zero for any psychological characteristic, positioning genetic effects as the first "law" of the field.[2] Heritability typically ranges from 30% to 60% for behavioral traits, with specific domains showing moderate to high values: approximately 50% for general cognitive ability (95% CI: 45-55%), 30-50% for personality dimensions, and 40-80% for psychiatric disorders like schizophrenia.[2][2] These patterns hold across diverse populations, from Western to Asian and Indian samples, indicating robustness beyond cultural confounds.[2]Genome-wide complex trait analysis and polygenic scores provide converging evidence, capturing 20-40% of twin-study heritability for many behavioral outcomes through common SNPs, though "missing heritability" persists due to rare variants and non-additive effects.[2] Environmental influences, predominantly non-shared (unique experiences plus measurement error), explain the bulk of the remaining variance, typically 40-60%, while shared environment effects diminish after childhood for most traits.[2] This distribution underscores that genetic factors do not determine behavior in isolation but interact with stochastic and individual-specific environments to produce observed variation. The absence of traits wholly free from genetic input challenges nurture-only models, emphasizing multifactorial causation grounded in empirical variance partitioning.[2]
Cognitive Abilities and Intelligence
Twin studies provide robust evidence for genetic influences on intelligence, typically measured via IQ tests or general cognitive ability (g). Monozygotic twins reared together exhibit intraclass correlations of approximately 0.85 for IQ, compared to 0.60 for dizygotic twins, implying broad-sense heritability (h2) estimates of 50-60% after accounting for shared environment.[108] These estimates rise with age, from around 40% in early childhood to 70-80% in adulthood, as indicated by longitudinal meta-analyses of over 11,000 twin pairs showing developmental increases in genetic variance.[109] Adoption studies corroborate this, with IQ correlations between unrelated adoptive siblings near zero, while biological relatives show persistent similarity regardless of rearing environment.[108]Heritability applies similarly across specific cognitive domains—such as verbal, spatial, and memory abilities—but g accounts for 40-50% of variance in these, with genetic correlations exceeding 0.70 between diverse cognitive tests, underscoring a common genetic architecture dominated by polygenic effects on general processing efficiency.[4] Genome-wide association studies (GWAS) have identified thousands of variants associated with intelligence, with polygenic scores derived from large-scale meta-analyses (e.g., N > 3 million for educational attainment as a proxy) explaining 4-10% of phenotypic variance in independent IQ samples, a figure that has doubled approximately every five years due to increasing sample sizes.[108] SNP-based heritability estimates from these genomic data suggest genetics could explain 20-30% of IQ variance, bridging the gap toward twin-study figures and confirming polygenicity involving many small-effect loci rather than rare high-impact mutations.[108]These findings hold across populations and socioeconomic strata, though heritability appears slightly lower in low-SES environments in some U.S. samples, potentially reflecting genotype-environment interaction where genetic potential is more constrained by adversity; however, this moderation is small and not universally replicated, with shared environment effects minimal (c2 < 10%) in high-SES or adult cohorts.[109] Non-shared environmental influences dominate remaining variance, emphasizing that while genes set a probabilistic range, unique experiences shape outcomes within it. Overall, cognitive abilities exemplify the pervasiveness of genetic influence in behavioral genetics, with molecular methods increasingly validating classical estimates.[4]
Personality and Temperament
Twin studies consistently indicate that genetic factors account for approximately 40-60% of the variance in Big Five personality traits—openness to experience, conscientiousness, extraversion, agreeableness, and neuroticism—with the remainder attributed primarily to non-shared environmental influences rather than shared family environments.[110][111] Meta-analyses of behavior genetic studies confirm these moderate heritability estimates (h² ≈ 0.40) across traits, showing limited moderation by factors such as age, sex, or measurement method, though slight variations exist; for instance, extraversion and neuroticism often exhibit heritability around 0.48, while agreeableness is somewhat lower at 0.35-0.42.[112][113] Family and adoption studies corroborate these findings, revealing that monozygotic twins reared apart display personality correlations comparable to those reared together, underscoring the causal role of genetics over shared rearing effects.[114]Temperament, often conceptualized as the early-emerging, biologically rooted core of personality encompassing dimensions like negative emotionality, positive affectivity, and inhibitory control, similarly demonstrates moderate genetic heritability in longitudinal twin designs, with estimates ranging from 20-50% during infancy and early childhood, increasing to align with adult personality levels by adolescence.[115][116] Behavioral genetic research highlights temperament's polygenic basis, with genome-wide association studies identifying associations with hundreds of genetic loci influencing neural pathways related to emotional regulation and arousal, though effect sizes per variant remain small.[36] Unlike personality facets, temperament shows greater stability in heritability across development, with genetic influences on traits like effortful control emerging as early as 6-12 months and persisting, driven by additive genetic variance rather than dominance or epistasis.[117]Empirical evidence from large-scale twin registries, such as those in Finland and Australia, further supports the pervasiveness of genetic effects, where intraclass correlations for temperament dimensions in monozygotic twins exceed dizygotic pairs by roughly double, implying h² > 0.30 even after accounting for rater bias.[118] These patterns hold across cultures and measurement instruments, including parent and self-reports, with minimal evidence for gene-environment correlations inflating estimates in non-Western samples.[119] Recent polygenic score analyses predict up to 5-10% of temperament variance, validating twin-derived heritabilities through molecular evidence while revealing overlaps with psychopathology liability.[120]
Behavioral Disorders and Psychopathology
Twin and family studies consistently indicate moderate to high heritability for most psychiatric disorders, reflecting pervasive genetic influences on liability to behavioral disorders and psychopathology. Heritability estimates, which quantify the proportion of phenotypic variance attributable to genetic factors, range from approximately 30% for major depressive disorder to over 80% for psychotic conditions like schizophrenia. These figures derive primarily from classical twin designs comparing monozygotic and dizygotic concordance, as well as extended family and sibling analyses, which control for shared environments to isolate genetic effects. Genome-wide association studies (GWAS) further corroborate these findings through SNP-based heritability (h²-SNP), though estimates are lower (10-30%) due to capturing only common variants and polygenic architecture.[121]Psychotic disorders exhibit among the highest genetic loadings. For schizophrenia under Research Diagnostic Criteria, the Maudsley Twin Register yielded a heritability of 82%, with similar estimates of 85% for schizoaffective disorder and 84% for mania (indicative of bipolar spectrum liability). Sibling-based national registry data report 57% heritability for schizophrenia, aligning with meta-analyses confirming its status as a complex trait under strong polygenic control. Bipolar disorder shows comparable familial aggregation, with sibling heritability around 54%, though twin studies often exceed 80%, underscoring additive genetic dominance.[122][121][122]Neurodevelopmental disorders like autism spectrum disorder (ASD) and attention-deficit/hyperactivity disorder (ADHD) also display robust genetic influences. Meta-analyses of twin studies estimate ASD heritability at 64-91%, with population-based cohorts reinforcing high familial risk driven by genetic factors rather than shared environment. For ADHD, the mean heritability across 37 twin studies is 74%, with longitudinal designs attributing stability to persistent genetic effects over development.[123][124][125]Internalizing disorders such as major depressive disorder (MDD) show moderate heritability, typically 30-50%, with twin estimates of 41-49% and sibling data (including half-siblings reared apart) at 36-51%, higher in cases with early onset or comorbidity. Obsessive-compulsive disorder and anorexia nervosa yield 38% and 41% sibling heritability, respectively, while alcohol dependence reaches 44%. Across disorders, non-shared environmental influences predominate over shared family effects, and genetic correlations (e.g., between schizophrenia and bipolar) suggest pleiotropy, where common variants contribute to multiple psychopathologies. These patterns hold after accounting for ascertainment biases, emphasizing causal genetic realism over purely environmental narratives.[126][121][121]
Environmental Contributions and Non-Shared Effects
In behavioral genetics, environmental variance in traits is decomposed into shared environmental effects (C), which systematically influence siblings or family members similarly (e.g., socioeconomic status or general family rearing practices), and non-shared environmental effects (E), which are unique to individuals and contribute to differences even among those raised together (e.g., differential sibling treatment, peer groups, illnesses, or measurement error).[127] This partition arises from the classical twin design, where monozygotic (MZ) twins reared together provide estimates of total genetic and shared environmental similarity, contrasted with dizygotic (DZ) twins to isolate additive genetics from shared environment.[128] Non-shared effects (E) also absorb non-systematic variance, including stochastic developmental noise and potential child-driven experiences where genotypes evoke unique environments.[129]Empirical estimates from meta-analyses of twin and adoption studies reveal that shared environmental effects (C) typically explain 0-10% of variance in adult personality traits, with heritability (A) around 40% and the balance (50-60%) ascribed to non-shared effects (E).[8] For intelligence, shared environment accounts for up to 20-30% of variance in childhood but declines to near zero by adolescence or adulthood, where heritability rises to 50-80% and non-shared effects comprise the residual 20-50%.[127] In psychopathology, such as externalizing behaviors, shared environment shows modest effects (10-20%) early in life but fades, leaving non-shared effects dominant alongside genetics.[130] These patterns hold across large-scale twin registries, including the Minnesota Study of Twins Reared Apart, where MZ twins separated early exhibited correlations akin to those reared together, underscoring minimal shared environmental impact.[131]Non-shared effects, while empirically substantial, prove elusive to pinpoint causally, as attempts to identify specific mechanisms—such as unique peer influences or parental differentials—often fail replication or reveal genetic confounds, where siblings' genotypes elicit divergent experiences.[129] Turkheimer and colleagues argue that non-shared variance includes irreducible error and unpredictable events, complicating intervention designs that assume malleable shared factors.[132] Nonetheless, recent longitudinal analyses confirm non-shared effects as real, albeit often random and non-systematic, driving individual differences without familial aggregation.[8] This emphasis shifts research toward idiographic models, recognizing that environmental contributions to behavior are predominantly personalized rather than communal.[133]
Stability and Developmental Changes in Heritability
Heritability estimates for many behavioral traits, derived primarily from twin and adoption studies, tend to increase across development, reflecting a growing proportion of phenotypic variance attributable to genetic factors as individuals age. This pattern is attributed to diminishing influences of shared environments in childhood, which constrain genetic expression early on, and the amplification of genetic effects through mechanisms such as active gene-environment correlation, where individuals increasingly select environments congruent with their genotypes.[109][134]For cognitive abilities, particularly general intelligence (g), longitudinal twin data demonstrate a linear rise in heritability from approximately 41% at age 9 years to 55% at age 12 years and 66% by ages 16-20 years.[135] This upward trajectory, termed the Wilson effect, plateaus at around 80% by ages 18-20 and remains stable into later adulthood, with early estimates as low as 20% in infancy giving way to mature levels exceeding 70-80%.[134][109] Concurrently, shared environmental contributions, prominent in early childhood (accounting for up to 40-50% of variance), largely dissipate by adolescence, shifting variance toward nonshared environments and genetics.[135]In personality traits, such as extraversion and neuroticism, heritability shows moderate to high stability over time, with genetic factors explaining continuity from adolescence into adulthood; for instance, twin studies report heritability estimates rising or stabilizing around 40-50% for these dimensions in longitudinal assessments.[136] For psychopathology, patterns vary: heritability of externalizing behaviors and overall problem scores increases across adolescence, while internalizing traits like anxiety exhibit more stable or modest genetic influences (~20-50%).[137][138]Borderline personality disorder traits, for example, display heightened heritability in adulthood compared to adolescence, aligning with broader trends in behavioral disorders.[138]A meta-analysis of longitudinal behavioral genetic studies across eight phenotype domains (including IQ, personality, and problem behaviors) confirms age-related heritability increases for most traits from adolescence to young adulthood, driven by persistent genetic effects amid fading shared environment impacts.[137] Age-to-age phenotypic stability in these traits is predominantly genetic, with correlations between occasions largely mediated by shared additive genetic variance rather than environmental persistence.[109] These developmental shifts underscore the dynamic interplay of genetic and environmental variances, where early plasticity yields to genetically influenced canalization in later stages.[139]
Underlying Mechanisms
Polygenicity and Effect Sizes
Behavioral traits, including cognitive abilities, personality dimensions, and psychopathology, display a highly polygenic genetic architecture, characterized by the involvement of thousands to millions of genetic variants—predominantly common single nucleotide polymorphisms (SNPs)—each exerting small incremental effects on phenotypic outcomes.[140][141] This distributed influence has been empirically demonstrated through genome-wide association studies (GWAS), which systematically test associations between SNPs and traits across the genome, revealing no single variant or small set of loci accounting for substantial variance.[142] Instead, polygenic scores (PGS), which aggregate weighted effects from vast numbers of SNPs (often exceeding 1 million genome-wide), provide the primary means to quantify cumulative genetic propensity, capturing 5–15% of trait variance in independent samples for complex behaviors.[38][143]For intelligence and cognitive performance, GWAS meta-analyses have identified hundreds of independent loci; a 2023 study of 78,308 individuals reported novel associations, adding to prior discoveries of over 200 loci in larger cohorts, with ongoing increases as sample sizes expand.[144] Individual SNP effect sizes remain minuscule, typically explaining less than 0.05% of phenotypic variance per variant, consistent with models where rarer alleles may carry slightly larger but still modest effects, and common variants dominate due to their prevalence.[145][146] These small per-locus contributions necessitate massive sample sizes (often hundreds of thousands) for detection, underscoring why earlier candidate gene studies, focusing on presumed large-effect variants, largely failed to replicate.[147]Personality traits exhibit analogous polygenicity, with GWAS implicating numerous loci across dimensions like neuroticism, extraversion, and conscientiousness; for example, a 2024 analysis identified 254 genes associated with at least one Big Five trait, derived from variants with individually negligible impacts.[36] Effect sizes per SNP for personality are comparably tiny, often below 0.01% of variance, reflecting a polygenic and poly-environmental etiology where genetic influences interweave with non-shared environmental factors.[114] PGS for personality, summing effects from thousands of SNPs, predict modest portions of trait variance (e.g., 1–5%) and correlate with related outcomes like life satisfaction, but their limited explanatory power highlights the challenge of infinitesimal effects distributed across the genome.[148]In behavioral disorders and psychopathology, polygenicity is similarly pervasive, with traits like externalizing behaviors involving PGS from approximately 1 million SNPs, each with effect sizes too small for standalone significance yet aggregating to explain behavioral deviations.[143] Regional polygenicity estimates from GWAS summary statistics suggest thousands of causal variants per trait, supporting causal realism in which behavioral outcomes arise from myriad subtle genetic perturbations rather than deterministic major genes.[149] This architecture implies that future advances in PGS accuracy will depend on even larger, diverse cohorts to refine effect size estimates and mitigate biases from European-ancestry data dominance.[150]
Gene-Environment Interplay and Epigenetics
Gene-environment correlations (rGE) occur when genetic differences among individuals are associated with differences in their environments, influencing behavioral outcomes. These include passive rGE, where parents transmit both genetic predispositions and rearing environments to offspring; evocative rGE, in which an individual's genotype elicits responses from others that shape the environment; and active rGE, where genetically influenced traits lead individuals to seek or create compatible environments.[151] Twin and adoption studies demonstrate that rGE contributes substantially to variance in traits like externalizing behaviors, with genetic factors driving selection into peer groups or family dynamics that amplify initial propensities.[152] For instance, children with heritable antisocial tendencies often evoke harsher parenting or affiliate with deviant peers, perpetuating cycles of maladaptation.[153]Gene-environment interactions (GxE) represent instances where environmental influences on a phenotype vary by genotype, often modeled as the product term in variance decompositions such as Var(P)=g+ϵ+g×ϵ.[154] In behavioral genetics, twin studies detect GxE by comparing monozygotic and dizygotic concordance across environmental strata, revealing amplified genetic effects in supportive settings or heightened environmental sensitivity for high-risk genotypes.[155] A classic example is the interaction between childhood maltreatment and variants in the MAOA gene, where low-activity alleles predict increased aggression risk under adversity, though replication has been inconsistent in polygenic contexts.[156] Polygenic scores for educational attainment interact with socioeconomic status, such that genetic advantages yield greater cognitive gains in enriched environments, underscoring non-additive dynamics.[157] These effects, while statistically significant, typically explain modest additional variance beyond main effects, challenging claims of dominant interplay.[158]Epigenetics provides a molecular mechanism for GxE, involving heritable changes in gene expression without DNA sequence alterations, such as DNA methylation or histone modifications responsive to environmental cues.[159] In behavioral contexts, adverse early experiences like maternal deprivation in rodents induce epigenetic marks on stress-related genes (e.g., glucocorticoid receptor), altering offspring behavior and stress reactivity, with some effects persisting transgenerationally in animal models.[159] Human studies link prenatal famine exposure to methylation changes correlating with later metabolic and cognitive traits, suggesting environment-induced epigenetics mediates long-term outcomes.[160] However, epigenetic contributions to behavioral heritability remain small and unstable, with low inter-individual reliability of marks and limited evidence for causal roles in complex traits like personality, as twin discordance often exceeds epigenetic variance.[161] Claims of epigenetics resolving "missing heritability" lack robust support in humans, where genetic sequence variants predominate.[160]
Neurobiological Pathways from Genes to Behavior
Genetic variants exert their influence on behavior primarily through alterations in brainstructure, neural connectivity, and neurochemical signaling, mediated by molecular processes such as gene expressionregulation and protein synthesis during neurodevelopment. Twin studies indicate that heritability estimates for brain volumes range from 40% to 90% across the lifespan, with subcortical structures showing particularly high genetic control (44-88%) and cortical regions exhibiting moderate to high heritability that increases with age.[162][163] These structural differences, including variations in cortical thickness (SNP-based h² ≈ 0.25 globally, up to 0.28 in sensorimotor regions) and white matter hyperintensity volume (h² ≈ 0.22), correlate genetically with cognitive functions like executive control and memory.[164] For instance, polygenic scores for inferior frontal gyrus thickness predict enhanced executive function in the absence of cardiovascular comorbidities.[164]At the molecular level, genes encoding neurotransmitter systems form key pathways linking genotype to behavioral phenotypes. Dopaminergic genes, such as DRD4 and COMT, modulate reward processing and prefrontal dopamine availability, contributing to traits like extraversion and impulsivity, with effects amplified in high-stress environments.[165] Serotonergic polymorphisms in SLC6A4 and HTR2A influence anxiety-related behaviors by altering serotonin reuptake and receptor density in limbic regions, with meta-analyses confirming associations across personality inventories.[166] These variants affect synaptic plasticity and circuit formation during critical developmental windows, where even common alleles disrupt neuronal migration or pruning, elevating risk for psychopathology such as ADHD or schizophrenia.[167]Functional neuroimaging reveals genetic impacts on resting-state connectivity and task-evoked activity, bridging structural changes to observable behaviors. For example, GWAS-identified loci for global cortical thickness overlap with expression in prefrontal and cingulate cortices, regions implicated in decision-making and emotional regulation, yielding genetic correlations with intelligence (rg ≈ -0.2 for white matter hyperintensities) and psychiatric traits like insomnia.[164] In social behavior, orthologous genes like AVPR1A (vasopressin receptor) regulate hypothalamic circuits for affiliation and aggression, as evidenced by sequence variations altering pair-bonding in voles and paralleling human attachment styles.[168] Overall, these polygenic pathways underscore a causal chain from DNA sequence to neural architecture to adaptive or maladaptive behaviors, with small-effect variants cumulatively shaping individual differences.[169]
Controversies and Ideological Debates
Scientific Limitations and Methodological Critiques
Classical twin studies, foundational to behavioral genetics, rely on the equal environments assumption, positing that monozygotic (MZ) and dizygotic (DZ) twins experience equivalently similar trait-relevant environments, such that differences in trait similarity reflect genetic factors rather than environmental ones.[170] Violations of this assumption, where MZ twins encounter more similar environments due to greater parental treatment similarity or self-selection into alike experiences, lead to overestimation of heritability; for instance, simulations demonstrate that such violations can inflate heritability estimates by up to 20-30% for behavioral traits like political attitudes.[64] Empirical tests, including retrospective reports and co-twin control designs, confirm EEA breaches particularly for subjective traits such as personality and ideology, though less so for objective physical traits.[171]Heritability estimates from twin and family designs are further susceptible to biases from unmodeled factors like assortative mating, where phenotypic similarity between parents correlates with genetic similarity, artificially elevating observed twin correlations and confounding additive genetic variance.[43] Measurement error in phenotypic assessment attenuates heritability downward by inflating total variance without proportionally affecting genetic covariance, a ceiling effect documented in behavioral traits where rater or self-report inconsistencies reduce estimates by 10-20%; for example, teacher-reported ADHD shows twin heritability of 69% but requires disattenuation for reliable comparison to molecular estimates.[172] Participation and selection biases exacerbate this, as genetically influenced traits correlate with study inclusion, underestimating heritability for low-participating subgroups in large cohorts.[173]The "missing heritability" gap represents a core methodological challenge, wherein twin/family-based broad-sense heritability (often 40-80% for cognitive and personality traits) far exceeds SNP-based narrow-sense heritability from genome-wide association studies (GWAS), typically capturing only 10-30% of variance for complex behaviors.[174] This discrepancy arises partly from twin overestimation due to EEA violations and unaccounted epistasis, where genetic interactions generate "phantom heritability" inflating classical estimates; models of limiting pathway epistasis, applicable to polygenic traits, show true heritability reduced by 50% or more relative to apparent values.[175] Rare variants, structural variants, and imperfect linkage disequilibrium further evade GWAS detection, contributing substantially to unresolved variance in behavioral polygenic scores.[174]Causal inference remains limited, as heritability quantifies variance partitioning within populations under specific conditions but does not delineate mechanisms or generalizability across environments; for instance, estimates vary by socioeconomic homogeneity, rising in uniform settings where gene expression is less moderated.[43] Replication challenges in GWAS for behavioral traits stem from small effect sizes (often <0.01% per SNP), polygenicity involving thousands of loci, and population stratification, yielding inconsistent hits across cohorts without functional validation.[176] Gene-environment interplay, including dynamic transactions where genotypes evoke environments, is inadequately modeled in additive frameworks, confoundingadditive genetic effects with interactive terms.[43] These issues underscore the need for integrative approaches, such as extended twin-family designs or whole-genome sequencing, to mitigate biases and enhance precision.[177]
Challenges with Heritability Estimates and Missing Variance
Heritability estimates in behavioral genetics, derived primarily from twin and family studies, typically attribute 30-60% of variance in traits such as personality dimensions or psychopathology to genetic factors, with higher figures (50-80%) for cognitive abilities like intelligence.[43] These broad-sense estimates encompass additive, dominance, and epistatic genetic effects but rely on key assumptions, including the equal environments assumption (EEA), which posits that monozygotic (MZ) and dizygotic (DZ) twins experience equivalently similar environments. Empirical tests across 32 outcomes, including personality and cognitive traits, indicate that EEA violations occur but introduce only modest bias, significantly reducing heritability estimates in few cases like neuroticism while supporting overall validity for most behavioral traits.[178]A prominent challenge arises from assortative mating, where individuals select partners based on phenotypic similarity for heritable traits like intelligence or extraversion, violating standard twin model assumptions of random mating. This phenomenon increases genetic variance across generations and can bias DZ twin correlations upward, leading to underestimated shared environmental effects or inflated heritability in unadjusted models; simulations and empirical data from cohorts like the UK Biobank confirm it downwardly biases SNP-based heritability estimates while complicating twin-derived figures without explicit modeling.[179][180] Additionally, twin estimates conflate genetic and prenatal environmental sharing in MZ twins, potentially overstating heritability for early-emerging behaviors, though adoption studies and reared-apart twin data largely corroborate findings after corrections.The "missing heritability" problem exacerbates these issues, revealing a persistent gap between twin/family estimates and those from genome-wide association studies (GWAS) or SNP heritability methods, which capture only common additive variants. For childhood behavioral problems, twin heritability averages 52% across parent, teacher, and self-reports, yet SNP heritability averages just 6%, representing only 11-13% of the twin figure; similar discrepancies appear in personality (twin ~40-50%, molecular ~10-20%) and intelligence (twin 50-80%, GWAS/polygenic scores ~10-15%).[181][182] This threefold gap—h²_GWAS < h²_SNP < h²_twin—stems from molecular methods missing rare variants, copy-number variations, non-additive interactions, and gene-environment interplay, which twin designs aggregate without distinguishing; behavioral traits, being highly polygenic with small effect sizes, amplify detection challenges in underpowered GWAS.[182][175]Unexplained or "missing" variance in these models often attributes the remainder to non-shared environments (unique experiences plus measurement error), comprising 40-50% even at high heritability, but the molecular gap underscores undetected genetic contributions rather than purely environmental ones. Critics argue twin overestimation from unmodeled factors like cultural transmission or rater bias in self-reported behaviors, yet converging evidence from extended family designs and genomic relatedness matrices affirms substantial genetic influence, suggesting the gap reflects methodological incompleteness in molecular approaches rather than invalidation of classical estimates. Heritability figures remain population- and context-specific, varying with socioeconomic conditions or age, limiting direct causal inferences without integrating multilevel data.[171][183]
Replication and Causal Inference Issues
Behavioral genetics has largely avoided the replication crisis afflicting much of psychological science, where only about 36% of studies replicate, thanks to the field's emphasis on large effect sizes (heritabilities of 30%-50% for most traits) and meta-analytic practices that incentivize robust variance component estimation.[2] Twin and adoption studies consistently replicate findings such as the genetic mediation of trait correlations (e.g., 53%-65% for cognitive abilities) and the increase in intelligence heritability from childhood (around 20%-40%) to adulthood (up to 66%).[2] These replications stem from the polygenic nature of traits, involving many genes of small effect, and the field's focus on quantitative extremes rather than rare variants.[2]Early molecular efforts, however, encountered severe replication issues, particularly in candidate gene studies targeting specific polymorphisms (e.g., for serotonin or dopamine systems in personality or psychopathology), which often failed in larger follow-ups due to inadequate power, population stratification, and selective reporting.[68][67] Over 1,000 such associations were proposed by 2012, but meta-analyses revealed most evaporated, with effect sizes inflated by small samples (often n<1,000) and p-hacking.[67] Genome-wide association studies (GWAS) mitigate this through stringent thresholds and massive cohorts (e.g., millions for educational attainment), yielding replicable hits, though they account for only 10%-20% of twin-study heritability, highlighting unresolved "missing heritability" from rare variants or non-additive effects.[2]Causal inference poses distinct challenges, as twin studies decompose variance but cannot directly establish directionality or rule out gene-environment correlations (rGE), where genotypes evoke environments (e.g., genetically impulsive children eliciting harsher parenting).[184] The equal environments assumption (EEA)—positing similar trait-relevant exposures for monozygotic (MZ) and dizygotic (DZ) twins—underpins heritability estimates but faces criticism for potential MZ-specific similarity (e.g., in treatment by parents), with simulations indicating up to 20%-30% heritability inflation if violated for psychosocial traits.[170][64] Empirical tests, including perceptions of twin similarity and virtual twin designs, largely support the EEA for cognitive and behavioral traits, though violations appear more pronounced for politically sensitive or culturally influenced outcomes.[185][170]Mendelian randomization (MR), leveraging genetic variants as instrumental variables, advances causal claims (e.g., genetically proxied education causally lowering BMI), but behavioral traits complicate this due to assortative mating, dynastic effects, and horizontal pleiotropy, where variants influence outcomes via unmeasured paths.[103][74] Weak instruments (low variance explained, often <5%) and population stratification bias estimates, particularly for socio-behavioral exposures like smoking or cognition, requiring sensitivity analyses like MR-Egger, yet false positives persist without replication in independent samples.[186][74] Family-based designs and natural experiments (e.g., policy discontinuities) complement MR by addressing rGE and stratification, but all methods demand large, diverse cohorts to disentangle polygenic causation from confounding.[184]
Sociopolitical Objections and Responses
Critics of behavioral genetics have raised sociopolitical objections primarily centered on the perceived threat to egalitarian ideals and individual agency, arguing that genetic influences imply fixed hierarchies incompatible with social justice frameworks. These concerns often invoke the determinism fallacy, misinterpreting heritability as predestining behavior without environmental input or change, which purportedly undermines efforts for societal reform.[187] Such views align with blank slate ideology, which denies substantial innate variation to prioritize nurture-based explanations for disparities, as articulated in critiques dismissing twin study evidence as methodologically flawed or ideologically driven.[11]Responses counter that heritability quantifies population-level variance attributable to genetic factors under specific environments, not individual-level causation or immutability; for instance, traits with high heritability like intelligence (h² ≈ 0.5–0.8 in adulthood) remain responsive to interventions, as demonstrated by environmental enrichments altering outcomes in adoption studies.[47] Meta-analyses of over 17,000 traits from twin studies confirm average heritability of 49% for behavioral phenotypes, including personality and cognition, refuting strict blank slatism by showing consistent genetic effects across diverse populations and rearing conditions.[47] This probabilistic model, incorporating gene-environment interactions, preserves causal realism without negating free will or policy efficacy, with proponents noting that ideological resistance in academia—evident in selective rejection of heritability data—stems from commitments to environmental determinism rather than empirical scrutiny.[188]Objections extend to fears of reviving eugenics through genetic insights into group differences, such as observed IQ gaps between racial or ethnic populations (e.g., 10–15 points between U.S. Black and White averages persisting since the 1970s), which critics claim could rationalize discriminatory policies or overlook systemic inequities.[189] Historical eugenics movements in the early 20th century misused nascent genetics for coercive sterilization of over 60,000 individuals in the U.S., fueling accusations that modern polygenic scores revive racial supremacist pseudoscience.[10] Policy implications are contested, with detractors warning against tailoring education or welfare based on heritable traits, lest it entrench inequality.Defenders respond that behavioral genetics repudiates eugenics' deterministic and coercive ethos, emphasizing voluntary, individual-level applications like personalized interventions rather than population control; organizations like the Behavior Genetics Association explicitly denounce misuse for group-based discrimination or genetic fatalism.[12] Within-group heritability, consistently moderate-to-high across White, Black, and Hispanic samples (h² ≈ 0.6–0.8), does not causally explain between-group variances, which likely involve gene-environment covariances and cultural factors, though admixture and GWAS data suggest partial genetic contributions in some cases without warranting group-targeted policies.[190][191] Free inquiry into such differences, as argued in reviews, avoids unintended harms like ignoring modifiable environmental levers, prioritizing evidence-based individualism over ideologically motivated suppression, which has historically biased mainstream discourse against genetic realism.[188]
Determinism Fallacy and Blank Slate Ideology
The determinism fallacy in behavioral genetics refers to the misconception that evidence of genetic influences on traits implies strict predetermination, eliminating scope for environmental modification, individual agency, or developmental plasticity. Heritability estimates, which quantify the proportion of phenotypic variance attributable to genetic variance within a specific population and environment, do not equate to fixed causal determinism for individuals; a trait with high heritability, such as height (h² ≈ 0.80 in well-nourished populations), can still respond to environmental changes like improved nutrition, as evidenced by secular increases in average stature over the 20th century. Similarly, for behavioral traits like IQ (h² ≈ 0.50-0.80 in adulthood), twin studies reveal that while genetic factors explain much of the variance, interventions such as enriched early education can yield lasting gains, underscoring gene-environment interplay rather than inevitability.[9][43][192]Critics of behavioral genetics often invoke this fallacy to dismiss heritability findings as endorsing fatalism, yet empirical data from adoption and twin rearing studies consistently demonstrate that genetic predispositions are probabilistic and context-dependent, not rigidly prescriptive; for instance, identical twins reared apart exhibit correlations lower than monozygotic concordance for many traits, reflecting non-shared environmental influences. This misinterpretation persists partly due to conflating population-level statistics with personal outcomes, ignoring that heritability rises with environmental uniformity—paradoxically, homogeneous environments amplify apparent genetic effects while masking malleability.[43][193]The blank slate ideology, rooted in empiricist traditions positing the human mind as a tabula rasa devoid of innate structures beyond basic sensory capacities, posits that behavioral differences arise overwhelmingly from cultural, social, and experiential factors. This view, historically championed in psychology by figures like B.F. Skinner who emphasized operant conditioning over endowment, has been empirically refuted by behavioral genetic evidence showing moderate to high heritability for complex traits including personality dimensions (h² ≈ 0.40-0.60), educational attainment (h² ≈ 0.40), and even ideological leanings (h² ≈ 0.60 in large twin cohorts). Large-scale genome-wide association studies further identify polygenic scores predicting behavioral outcomes, contradicting pure environmental determinism.[194][195][196]Despite such data, blank slate assumptions endure in academic and media discourse, often selectively applied to downplay innate sex differences or group variations while accepting genetic explanations for ideologically congenial traits, a pattern attributable to systemic biases favoring egalitarian narratives over causal evidence. This ideological commitment impedes policy realism, as interventions assuming infinite environmental malleability—such as certain educational reforms—underperform when genetic baselines are unaddressed, whereas acknowledging heritability informs targeted strategies like personalized learning. Primary sources from twin registries, less prone to publication biases in ideologically charged fields, affirm genetic contributions without negating nurture's role.[197][196]
Group Differences, Policy Implications, and Eugenics Fears
Observed differences in cognitive abilities, such as IQ, persist between racial and ethnic groups, with meta-analyses reporting averages of approximately 106 for East Asians, 100 for Europeans, and 85 for African Americans in the United States.[198] These gaps have narrowed modestly over decades but remain substantial, correlating with socioeconomic outcomes like educational attainment and income.[198] Twin and adoption studies indicate high within-group heritability for IQ (around 0.5-0.8 across groups), but between-group differences cannot be fully explained by environmental factors alone, as evidenced by transracial adoptions where Black children raised in White families still average IQs closer to racial norms than adoptive family means.[190][198]Genome-wide association studies (GWAS) and polygenic scores provide converging evidence for a partial genetic basis. Polygenic scores derived from educational attainment and intelligence GWAS predict cognitive performance across ancestries, with East Asians and Europeans showing higher scores than Africans, aligning with observed IQ hierarchies even after controlling for spatial autocorrelation in allele frequencies. These scores explain up to 10-15% of variance in IQ within populations and correlate with national IQ differences, supporting evolutionary selection pressures on cognitive traits.[199] Critics argue that portability issues across ancestries inflate environmental confoundings, yet replication in independent datasets and admixture studies (e.g., African Americans with varying European ancestry showing IQ gradients) bolster genetic interpretations.[200][198]Such findings challenge policies predicated on environmental equalization alone, like affirmative action or uniform educational standards, which assume malleable potentials leading to equal outcomes. Behavioral genetics suggests innate group variances imply that interventions should prioritize individual merit and targeted supports rather than quotas, as equalizing environments yields persistent gaps due to genetic limits on mean potentials.[198] For instance, precision education informed by genetic profiles could allocate resources efficiently, avoiding one-size-fits-all approaches that overlook heritability in learning abilities.[201] Policy resistance often stems from egalitarian ideologies fearing validation of hierarchies, yet ignoring genetics risks inefficient allocations, such as over-investing in closing unclosable gaps.[202]Eugenics fears arise from historical coercive programs, like early 20th-century sterilizations justified by hereditarian views, but modern behavioral genetics revives concerns over voluntary applications like embryo selection via polygenic screening. Advances in IVF and CRISPR enable selecting embryos for higher IQ polygenic scores, potentially boosting population means by 5-10 points per generation without state intervention.[11] Detractors equate this to "new eugenics," warning of designer babies exacerbating inequalities, yet proponents note it empowers parental choice, akin to selecting against diseases like Tay-Sachs, and counters dysgenic trends from differential fertility (lower-IQ groups having higher birth rates).[203][198] These fears, amplified in academia despite evidence of genetic influences, often serve to stigmatize research rather than engage ethical trade-offs, such as balancing autonomy against slippery slopes to coercion.[11] Empirical data from animal breeding and human selection experiments affirm feasibility without invoking historical atrocities.[203]
Applications and Future Directions
Implications for Education, Policy, and Intervention
Behavioral genetic research indicates that genetic factors account for approximately 50-60% of the variance in educational attainment, with heritability estimates increasing from childhood to adulthood due to active gene-environment correlations where individuals seek environments matching their genetic predispositions.[204][205] This suggests that educational systems assuming uniform environmental malleability may underperform, as polygenic scores derived from genome-wide association studies predict 10-15% of variance in years of schooling and cognitive skills, often outperforming socioeconomic status measures in forecasting outcomes.[206] Implications for education include shifting toward precision approaches, such as tailoring curricula to genetic propensities for traits like reading proficiency or perseverance, evidenced by studies showing genetic nurture effects where parental genotypes influence child environments via resource allocation.[207] Uniform interventions like broad early childhood programs show limited long-term gains partly because they overlook these genetic baselines, prompting recommendations for stratified tracking or individualized support to amplify genetic potential rather than enforcing equality of input.[208]In policy domains, high heritability of behavioral traits like intelligence and conscientiousness—estimated at 40-80%—challenges assumptions of outcome equality through redistribution alone, as genetic differences explain substantial portions of socioeconomic stratification beyond shared environment.[209] Policymakers incorporating behavioral genetics could prioritize meritocratic selection in higher education and employment, where genetic influences on achievement persist across contexts, reducing inefficiencies from quotas ignoring causal genetic variance.[53] For instance, cross-national comparisons reveal stronger genetic effects on attainment in merit-based systems like those in Scandinavia versus more equalizing U.S. environments, suggesting policies fostering opportunity equality—such as universal access to quality schooling—maximize societal returns without denying heritable limits on individual ceilings.[209] This contrasts with critiques of genetic fatalism, as heritability estimates do not preclude environmental modulation; rather, they inform realistic expectations, avoiding overinvestment in universal uplift programs with diminishing returns post-adolescence.[11]Interventions informed by behavioral genetics emphasize targeting gene-environment interactions, such as using polygenic risk scores to identify at-risk youth for enriched cognitive stimulation, where gains are larger for those with moderate genetic liabilities due to compensatory mechanisms.[210] Empirical data from twin studies support selective early interventions amplifying additive genetic variance, as shared environments explain less than 20% of attainment differences in adulthood, while nonshared experiences—potentially manipulable—interact with genotypes.[211] Advances in pharmacogenomics could extend to behavioral traits, with trials exploring genotype-specific responses to nootropics or ADHD medications showing heritability modulating efficacy, though broad application remains limited by polygenic complexity and ethical constraints on germline editing.[212] Overall, such strategies prioritize evidence-based allocation over ideologically driven universality, acknowledging that while environments can shift phenotypic expression within genetic bounds, population-level variance persists.[202]
Advances in Precision Genomics and Ethical Considerations
Recent advancements in genome-wide association studies (GWAS) have identified thousands of genetic variants associated with behavioral traits, enabling the construction of polygenic scores (PGS) that predict individual differences in intelligence, personality, and psychopathology. For instance, a 2024 GWAS of the 'big five' personality traits in approximately 224,000 individuals revealed hundreds of loci influencing traits like neuroticism and extraversion, with PGS explaining up to 5-10% of variance in these outcomes. Similarly, the Million Veteran Program's analysis of 635,969 participants in 2024 yielded genome-wide associations for 2,068 traits, including behavioral ones such as impulsivity and risk-taking, demonstrating the scalability of large cohorts in uncovering polygenic architecture. These developments build on earlier work, where PGS for educational attainment and cognitive ability now account for 10-15% of phenotypic variance in independent samples, surpassing prior estimates through improved statistical methods and diverse ancestries.[36][213]Precision genomics extends these findings toward causal inference and intervention, integrating PGS with functional genomics tools like CRISPR for validating variants' effects on neural pathways underlying behavior. A 2025 GWAS on delay discounting and impulsivity in 5,548 children identified novel loci, highlighting developmental stability of genetic influences from childhood onward. Projections indicate that by the mid-2030s, PGS for psychiatric and cognitive traits could predict over 10% of variance, facilitating personalized risk stratification for conditions like schizophrenia or ADHD. However, transferability remains limited across ancestries due to linkage disequilibrium differences, with European-ancestry GWAS dominating discovery samples, though multi-ancestry meta-analyses are mitigating this gap.[214][5][84]Ethical considerations in behavioral genomics center on balancing scientific progress with risks of misuse, emphasizing empirical scrutiny over ideological constraints. PGS applications raise privacy concerns, as genetic data aggregation in biobanks could enable unauthorized profiling or discrimination in employment and insurance, prompting calls for robust consent frameworks and data anonymization. Critics highlight potential exacerbation of social inequalities if PGS inform reproductive decisions like embryo selection via preimplantation genetic testing, yet proponents argue that denying access to predictive tools perpetuates unexamined environmental biases.[215][216][217]Methodological limitations, such as PGS explaining only a fraction of heritability and vulnerability to environmental confounders, necessitate caution against deterministic interpretations; ethical deployment requires transparent communication of probabilistic predictions rather than absolutes. Sociogenomic PGS for traits like educational attainment evoke eugenics fears, but evidence-based responses stress that genetic insights can inform targeted interventions, such as early cognitive enrichment, without endorsing coercion. Regulatory gaps persist, with legal uncertainties around PGS patenting and liability, underscoring the need for interdisciplinary oversight to prioritize causal evidence over precautionary suppression.[218][219][220]