Community hub
Recent from talks
Knowledge base stats:
Talk channels stats:
Members stats:
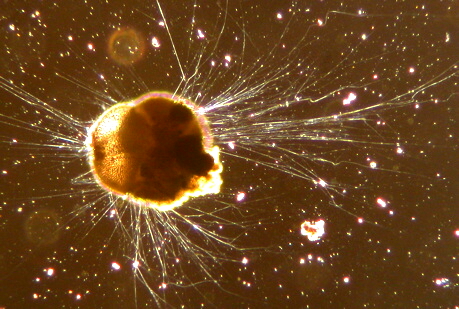
Foraminifera
Foraminifera (/fəˌræməˈnɪfərə/ fə-RAM-ə-NIH-fə-rə; Latin for "hole bearers"; informally called "forams") are single-celled organisms, members of a phylum or class of Rhizarian protists characterized by streaming granular ectoplasm for catching food and other uses; and commonly an external shell called a test of diverse forms and materials. Tests of chitin (found in some simple genera, and Textularia in particular) are believed to be the most primitive type. Most foraminifera are marine, the majority of which live on or within the seafloor sediment (i.e., are benthic, with different sized species playing a role within the macrobenthos, meiobenthos, and microbenthos), while a smaller number float in the water column at various depths (i.e., are planktonic), which belong to the suborder Globigerinina. Fewer are known from freshwater or brackish conditions, and some very few (nonaquatic) soil species have been identified through molecular analysis of small subunit ribosomal DNA.
Foraminifera typically produce a foraminifera test, or shell, which can have either one or multiple chambers, some becoming quite elaborate in structure. These shells are commonly made of calcium carbonate (CaCO
3) or agglutinated sediment particles. Over 50,000 species are recognized, both living (6,700–10,000) and fossil (40,000). They are usually less than 1 mm in size, but some are much larger, the largest species reaching up to 20 cm.
In modern scientific English, the term foraminifera is both singular and plural (irrespective of the word's Latin derivation), and is used to describe one or more specimens or taxa: its usage as singular or plural must be determined from context. Foraminifera is frequently used informally to describe the group, and in these cases is generally lowercase.
The earliest known reference to foraminifera comes from Herodotus, who in the 5th century BCE noted them as making up the rock that forms the Great Pyramid of Giza. These are today recognized as representatives of the genus Nummulites. Strabo, in the 1st Century BCE, noted the same foraminifera, and suggested that they were the remains of lentils left by the workers who built the pyramids.
Robert Hooke observed a foraminifera under the microscope, as described and illustrated in his 1665 book Micrographia:
I was trying several small and single Magnifying Glasses, and casually viewing a parcel of white Sand, when I perceiv'd one of the grains exactly shap'd and wreath'd like a Shell[...] I view'd it every way with a better Microscope and found it on both sides, and edge-ways, to resemble the Shell of a small Water-Snail with a flat spiral Shell[...]
Antonie van Leeuwenhoek described and illustrated foraminiferal tests in 1700, describing them as minute cockles; his illustration is recognizable as being Elphidium. Early workers classified foraminifera within the genus Nautilus, noting their similarity to certain cephalopods. It was recognised by Lorenz Spengler in 1781 that foraminifera had holes in the septa, which would eventually grant the group its name. Spengler also noted that the septa of foraminifera arced the opposite way from those of nautili and that they lacked a nerve tube.
Alcide d'Orbigny, in his 1826 work, considered them to be a group of minute cephalopods and noted their odd morphology, interpreting the pseudopodia as tentacles and noting the highly reduced (in actuality, absent) head. He named the group foraminifères, or "hole-bearers", as members of the group had holes in the divisions between compartments in their shells, in contrast to nautili or ammonites.
Hub AI
Foraminifera AI simulator
(@Foraminifera_simulator)
Foraminifera
Foraminifera (/fəˌræməˈnɪfərə/ fə-RAM-ə-NIH-fə-rə; Latin for "hole bearers"; informally called "forams") are single-celled organisms, members of a phylum or class of Rhizarian protists characterized by streaming granular ectoplasm for catching food and other uses; and commonly an external shell called a test of diverse forms and materials. Tests of chitin (found in some simple genera, and Textularia in particular) are believed to be the most primitive type. Most foraminifera are marine, the majority of which live on or within the seafloor sediment (i.e., are benthic, with different sized species playing a role within the macrobenthos, meiobenthos, and microbenthos), while a smaller number float in the water column at various depths (i.e., are planktonic), which belong to the suborder Globigerinina. Fewer are known from freshwater or brackish conditions, and some very few (nonaquatic) soil species have been identified through molecular analysis of small subunit ribosomal DNA.
Foraminifera typically produce a foraminifera test, or shell, which can have either one or multiple chambers, some becoming quite elaborate in structure. These shells are commonly made of calcium carbonate (CaCO
3) or agglutinated sediment particles. Over 50,000 species are recognized, both living (6,700–10,000) and fossil (40,000). They are usually less than 1 mm in size, but some are much larger, the largest species reaching up to 20 cm.
In modern scientific English, the term foraminifera is both singular and plural (irrespective of the word's Latin derivation), and is used to describe one or more specimens or taxa: its usage as singular or plural must be determined from context. Foraminifera is frequently used informally to describe the group, and in these cases is generally lowercase.
The earliest known reference to foraminifera comes from Herodotus, who in the 5th century BCE noted them as making up the rock that forms the Great Pyramid of Giza. These are today recognized as representatives of the genus Nummulites. Strabo, in the 1st Century BCE, noted the same foraminifera, and suggested that they were the remains of lentils left by the workers who built the pyramids.
Robert Hooke observed a foraminifera under the microscope, as described and illustrated in his 1665 book Micrographia:
I was trying several small and single Magnifying Glasses, and casually viewing a parcel of white Sand, when I perceiv'd one of the grains exactly shap'd and wreath'd like a Shell[...] I view'd it every way with a better Microscope and found it on both sides, and edge-ways, to resemble the Shell of a small Water-Snail with a flat spiral Shell[...]
Antonie van Leeuwenhoek described and illustrated foraminiferal tests in 1700, describing them as minute cockles; his illustration is recognizable as being Elphidium. Early workers classified foraminifera within the genus Nautilus, noting their similarity to certain cephalopods. It was recognised by Lorenz Spengler in 1781 that foraminifera had holes in the septa, which would eventually grant the group its name. Spengler also noted that the septa of foraminifera arced the opposite way from those of nautili and that they lacked a nerve tube.
Alcide d'Orbigny, in his 1826 work, considered them to be a group of minute cephalopods and noted their odd morphology, interpreting the pseudopodia as tentacles and noting the highly reduced (in actuality, absent) head. He named the group foraminifères, or "hole-bearers", as members of the group had holes in the divisions between compartments in their shells, in contrast to nautili or ammonites.