
Community hub
Recent from talks
Contribute something
Nothing was collected or created yet.
Biological pump
View on Wikipedia

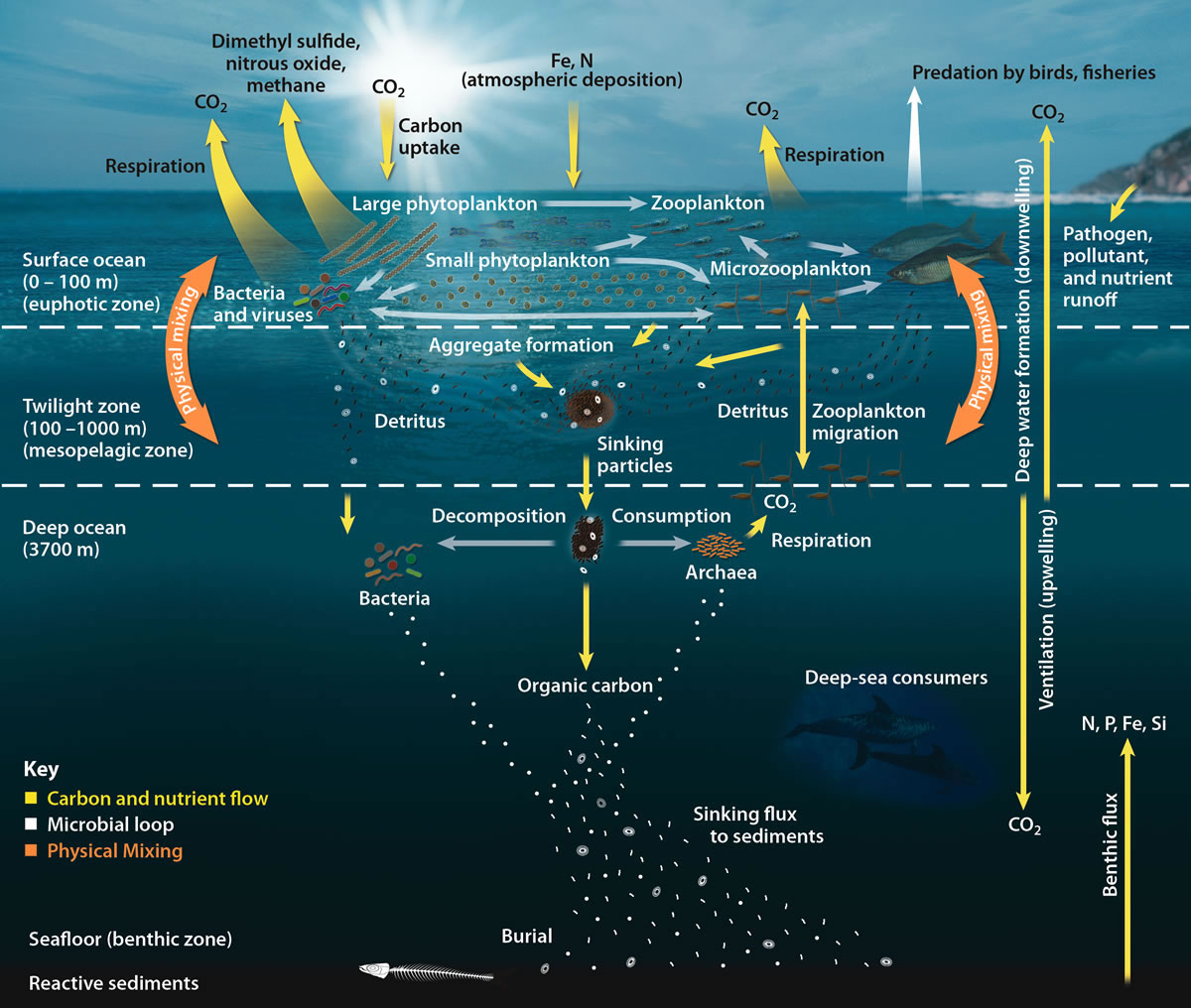
The biological pump (or marine biological carbon pump) is the ocean's biologically driven sequestration of carbon from the atmosphere and land runoff to the ocean interior and seafloor sediments.[1] In other words, it is a biologically mediated process which results in the sequestering of carbon in the deep ocean away from the atmosphere and the land. The biological pump is the biological component of the "marine carbon pump" which contains both a physical and biological component. It is the part of the broader oceanic carbon cycle responsible for the cycling of organic matter formed mainly by phytoplankton during photosynthesis (soft-tissue pump), as well as the cycling of calcium carbonate (CaCO3) formed into shells by certain organisms such as plankton and mollusks (carbonate pump).[2]
Budget calculations of the biological carbon pump are based on the ratio between sedimentation (carbon export to the ocean floor) and remineralization (release of carbon to the atmosphere).
The biological pump is not so much the result of a single process, but rather the sum of a number of processes each of which can influence biological pumping. Overall, the pump transfers about 10.2 gigatonnes of carbon every year into the ocean's interior and a total of 1300 gigatonnes carbon over an average 127 years.[3] This takes carbon out of contact with the atmosphere for several thousand years or longer. An ocean without a biological pump would result in atmospheric carbon dioxide levels about 400 ppm higher than the present day.
Overview
[edit]| Part of a series of overviews on |
| Marine life |
|---|
 |
The element carbon plays a central role in climate and life on Earth. It is capable of moving among and between the geosphere, cryosphere, atmosphere, biosphere and hydrosphere. This flow of carbon is referred to as the Earth's carbon cycle. It is also intimately linked to the cycling of other elements and compounds. The ocean plays a fundamental role in Earth's carbon cycle, helping to regulate atmospheric CO2 concentration. The biological pump is a set of processes that transfer organic carbon from the surface to the deep ocean, and is at the heart of the ocean carbon cycle.[4]
The biological pump depends on the fraction of primary produced organic matter that survives degradation in the euphotic zone and that is exported from surface water to the ocean interior, where it is mineralized to inorganic carbon, with the result that carbon is transported against the gradient of dissolved inorganic carbon (DIC) from the surface to the deep ocean. This transfer occurs through physical mixing and transport of dissolved and particulate organic carbon (POC), vertical migrations of organisms (zooplankton, fish) and through gravitational settling of particulate organic carbon.[5][6]: 526 [7]
The biological pump can be divided into three distinct phases, the first of which is the production of fixed carbon by planktonic phototrophs in the euphotic (sunlit) surface region of the ocean. In these surface waters, phytoplankton use carbon dioxide (CO2), nitrogen (N), phosphorus (P), and other trace elements (barium, iron, zinc, etc.) during photosynthesis to make carbohydrates, lipids, and proteins. Some plankton, (e.g. coccolithophores and foraminifera) combine calcium (Ca) and dissolved carbonates (carbonic acid and bicarbonate) to form a calcium carbonate (CaCO3) protective coating.[8]


| Part of a series on the |
| Carbon cycle |
|---|
 |
Once this carbon is fixed into soft or hard tissue, the organisms either stay in the euphotic zone to be recycled as part of the regenerative nutrient cycle or once they die, continue to the second phase of the biological pump and begin to sink to the ocean floor. The sinking particles will often form aggregates as they sink, which greatly increases the sinking rate. It is this aggregation that gives particles a better chance of escaping predation and decomposition in the water column and eventually making it to the sea floor.[8]
The fixed carbon that is decomposed by bacteria either on the way down or once on the sea floor then enters the final phase of the pump and is remineralized to be used again in primary production. The particles that escape these processes entirely are sequestered in the sediment and may remain there for millions of years. It is this sequestered carbon that is responsible for ultimately lowering atmospheric CO2.[8]
Biology, physics and gravity interact to pump organic carbon into the deep sea. The processes of fixation of inorganic carbon in organic matter during photosynthesis, its transformation by food web processes (trophodynamics), physical mixing, transport and gravitational settling are referred to collectively as the biological pump.[10]
The biological pump is responsible for transforming dissolved inorganic carbon (DIC) into organic biomass and pumping it in particulate or dissolved form into the deep ocean. Inorganic nutrients and carbon dioxide are fixed during photosynthesis by phytoplankton, which both release dissolved organic matter (DOM) and are consumed by herbivorous zooplankton. Larger zooplankton - such as copepods - egest fecal pellets which can be reingested and sink or collect with other organic detritus into larger, more-rapidly-sinking aggregates. DOM is partially consumed by bacteria (black dots) and respired; the remaining refractory DOM is advected and mixed into the deep sea. DOM and aggregates exported into the deep water are consumed and respired, thus returning organic carbon into the enormous deep ocean reservoir of DIC. About 1% of the particles leaving the surface ocean reach the seabed and are consumed, respired, or buried in the sediments. There, carbon is stored for millions of years. The net effect of these processes is to remove carbon in organic form from the surface and return it to DIC at greater depths, maintaining the surface-to-deep ocean gradient of DIC. Thermohaline circulation returns deep-ocean DIC to the atmosphere on millennial timescales.[10]
Primary production
[edit]
The first step in the biological pump is the synthesis of both organic and inorganic carbon compounds by phytoplankton in the uppermost, sunlit layers of the ocean.[13] Organic compounds in the form of sugars, carbohydrates, lipids, and proteins are synthesized during the process of photosynthesis:
CO2 + H2O + light → CH2O + O2
In addition to carbon, organic matter found in phytoplankton is composed of nitrogen, phosphorus and various trace metals. The ratio of carbon to nitrogen and phosphorus varies from place to place,[14] but has an average ratio near 106C:16N:1P, known as the Redfield ratio. Trace metals such as magnesium, cadmium, iron, calcium, barium and copper are orders of magnitude less prevalent in phytoplankton organic material, but necessary for certain metabolic processes and therefore can be limiting nutrients in photosynthesis due to their lower abundance in the water column.[8]
Oceanic primary production accounts for about half of the carbon fixation carried out on Earth. Approximately 50–60 Pg of carbon are fixed by marine phytoplankton each year despite the fact that they account for less than 1% of the total photosynthetic biomass on Earth. The majority of this carbon fixation (~80%) is carried out in the open ocean while the remaining amount occurs in the very productive upwelling regions of the ocean. Despite these productive regions producing 2 to 3 times as much fixed carbon per area, the open ocean accounts for greater than 90% of the ocean area and therefore is the larger contributor.[8]
Forms of carbon
[edit]
Dissolved and particulate carbon
[edit]Phytoplankton supports all life in the ocean as it converts inorganic compounds into organic constituents. This autotrophically produced biomass presents the foundation of the marine food web.[15] In the diagram immediately below, the arrows indicate the various production (arrowhead pointing toward DOM pool) and removal processes of DOM (arrowhead pointing away), while the dashed arrows represent dominant biological processes involved in the transfer of DOM. Due to these processes, the fraction of labile DOM decreases rapidly with depth, whereas the refractory character of the DOM pool considerably increases during its export to the deep ocean. DOM, dissolved organic matter.[15][16]


Ocean carbon pools
[edit]The marine biological pump depends on a number of key pools, components and processes that influence its functioning. There are four main pools of carbon in the ocean.[4]
- Dissolved inorganic carbon (DIC) is the largest pool. It constitutes around 38,000 Pg C [18] and includes dissolved carbon dioxide (CO2), bicarbonate (HCO−
3), carbonate (CO2−
3), and carbonic acid (H2CO3). The equilibrium between carbonic acid and carbonate determines the pH of the seawater. Carbon dioxide dissolves easily in water and its solubility is inversely related to temperature. Dissolved CO2 is taken up in the process of photosynthesis, and can reduce the partial pressure of CO2 in the seawater, favouring drawdown from the atmosphere. The reverse process respiration, releases CO2 back into the water, can increase partial pressure of CO2 in the seawater, favouring release back to the atmosphere. The formation of calcium carbonate by organisms such as coccolithophores has the effect of releasing CO2 into the water.[19][20][21][4] - Dissolved organic carbon (DOC) is the next largest pool at around 662 Pg C.[22] DOC can be classified according to its reactivity as refractory, semi-labile or labile. The labile pool constitutes around 0.2 Pg C, is bioavailable, and has a high production rate (~ 15−25 Pg C y−1).[23] The refractory component is the biggest pool (~642 Pg C ± 32;[22] but has a very low turnover rate (0.043 Pg C y−1).[23] The turnover time for refractory DOC is thought to be greater than 1000 years.[24][25][4]
- Particulate organic carbon (POC) constitutes around 2.3 Pg C,[26][27] and is relatively small compared with DIC and DOC. Though small in size, this pool is highly dynamic, having the highest turnover rate of any organic carbon pool on the planet.[28] Driven by primary production, it produces around 50 Pg C y−1 globally.[29][30][31] It can be separated into living (e.g. phytoplankton, zooplankton, bacteria) and non-living (e.g. detritus) material. Of these, the phytoplankton carbon is particularly important, because of its role in marine primary production, and also because it serves as the food resource for all the larger organisms in the pelagic ecosystem.[4]
- Particulate inorganic carbon (PIC) is the smallest of the pools at around 0.03 Pg C.[32] It is present in the form of calcium carbonate (CaCO3) in particulate form, and impacts the carbonate system and pH of the seawater. Estimates for PIC production are in the region of 0.8–1.4 Pg C y−1, with at least 65% of it being dissolved in the upper water column, the rest contributing to deep sediments.[33] Coccolithophores and foraminifera are estimated to be the dominant sources of PIC in the open ocean.[34][33] The PIC pool is of particular importance due to its role in the ocean carbonate system, and in facilitating the export of carbon to the deep ocean through the carbonate pump, whereby PIC is exported out of the photic zone and deposited in the bottom sediments.[35][4]
Calcium carbonate
[edit]


of the plates of buried coccolithophores ( see below ↓ )
Particulate inorganic carbon (PIC) usually takes the form of calcium carbonate (CaCO3), and plays a key part in the ocean carbon cycle.[36] This biologically fixed carbon is used as a protective coating for many planktonic species (coccolithophores, foraminifera) as well as larger marine organisms (mollusk shells). Calcium carbonate is also excreted at high rates during osmoregulation by fish, and can form in whiting events.[37] While this form of carbon is not directly taken from the atmospheric budget, it is formed from dissolved forms of carbonate which are in equilibrium with CO2 and then responsible for removing this carbon via sequestration.[38]
CO2 + H2O → H2CO3 → H+ + HCO3−
Ca2+ + 2HCO3− → CaCO3 + CO2 + H2O
While this process does manage to fix a large amount of carbon, two units of alkalinity are sequestered for every unit of sequestered carbon.[2][39] The formation and sinking of CaCO3 therefore drives a surface to deep alkalinity gradient which serves to raise the pH of surface waters, shifting the speciation of dissolved carbon to raise the partial pressure of dissolved CO2 in surface waters, which actually raises atmospheric levels. In addition, the burial of CaCO3 in sediments serves to lower overall oceanic alkalinity, tending to raise pH and thereby atmospheric CO2 levels if not counterbalanced by the new input of alkalinity from weathering.[1] The portion of carbon that is permanently buried at the sea floor becomes part of the geologic record. Calcium carbonate often forms remarkable deposits that can then be raised onto land through tectonic motion as in the case with the White Cliffs of Dover in Southern England. These cliffs are made almost entirely of the plates of buried coccolithophores.[40]
Oceanic carbon cycle
[edit]
Three main processes (or pumps) that make up the marine carbon cycle bring atmospheric carbon dioxide (CO2) into the ocean interior and distribute it through the oceans. These three pumps are: (1) the solubility pump, (2) the carbonate pump, and (3) the biological pump. The total active pool of carbon at the Earth's surface for durations of less than 10,000 years is roughly 40,000 gigatons C (Gt C, a gigaton is one billion tons, or the weight of approximately 6 million blue whales), and about 95% (~38,000 Gt C) is stored in the ocean, mostly as dissolved inorganic carbon.[41][42] The speciation of dissolved inorganic carbon in the marine carbon cycle is a primary controller of acid-base chemistry in the oceans.
Solubility pump
[edit]
The biological pump is accompanied by a physico-chemical counterpart known as the solubility pump. This pump transports significant amounts of carbon in the form of dissolved inorganic carbon (DIC) from the ocean's surface to its interior. It involves physical and chemical processes only, and does not involve biological processes.[43]
The solubility pump is driven by the coincidence of two processes in the ocean:
- The solubility of carbon dioxide is a strong inverse function of seawater temperature (i.e. solubility is greater in cooler water)
- The thermohaline circulation is driven by the formation of deep water at high latitudes where seawater is usually cooler and denser
Since deep water (that is, seawater in the ocean's interior) is formed under the same surface conditions that promote carbon dioxide solubility, it contains a higher concentration of dissolved inorganic carbon than might be expected from average surface concentrations. Consequently, these two processes act together to pump carbon from the atmosphere into the ocean's interior. One consequence of this is that when deep water upwells in warmer, equatorial latitudes, it strongly outgasses carbon dioxide to the atmosphere because of the reduced solubility of the gas.[44]
Carbonate pump
[edit]The carbonate pump is sometimes referred to as the "hard tissue" component of the biological pump.[45] Some surface marine organisms, like coccolithophores, produce hard structures out of calcium carbonate, a form of particulate inorganic carbon, by fixing bicarbonate.[46] This fixation of DIC is an important part of the oceanic carbon cycle.
Ca2+ + 2 HCO3− → CaCO3 + CO2 + H2O
While the biological carbon pump fixes inorganic carbon (CO2) into particulate organic carbon in the form of sugar (C6H12O6), the carbonate pump fixes inorganic bicarbonate and causes a net release of CO2.[46] In this way, the carbonate pump could be termed the carbonate counter pump. It works counter to the biological pump by counteracting the CO2 flux into the biological pump.[47]
Continental shelf pump
[edit]The continental shelf pump is proposed as operating in the shallow waters of the continental shelves as a mechanism transporting carbon (dissolved or particulate) from the continental waters to the interior of the adjacent deep ocean.[48] As originally formulated, the pump is thought to occur where the solubility pump interacts with cooler, and therefore denser water from the shelf floor which feeds down the continental slope into the neighbouring deep ocean.[48] The shallowness of the continental shelf restricts the convection of cooling water, so the cooling can be greater for continental shelf waters than for neighbouring open ocean waters. These cooler waters promote the solubility pump and lead to an increased storage of dissolved inorganic carbon. This extra carbon storage is further augmented by the increased biological production characteristic of shelves.[49] The dense, carbon-rich shelf waters then sink to the shelf floor and enter the sub-surface layer of the open ocean via isopycnal mixing.[48] As the sea level rises in response to global warming, the surface area of the shelf seas will grow and in consequence the strength of the shelf sea pump should increase.[50]
Processes in the biological pump
[edit]
In the diagram on the right, phytoplankton convert CO2, which has dissolved from the atmosphere into the surface oceans (90 Gt yr−1), into particulate organic carbon (POC) during primary production (~ 50 Gt C yr−1). Phytoplankton are then consumed by copepods, krill and other small zooplankton grazers, which in turn are preyed upon by higher trophic levels. Any unconsumed phytoplankton form aggregates, and along with zooplankton faecal pellets, sink rapidly and are exported out of the mixed layer (< 12 Gt C yr−1 14). Krill, copepods, zooplankton and microbes intercept phytoplankton in the surface ocean and sinking detrital particles at depth, consuming and respiring this POC to CO2 (dissolved inorganic carbon, DIC), such that only a small proportion of surface-produced carbon sinks to the deep ocean (i.e., depths > 1000 m). As krill and smaller zooplankton feed, they also physically fragment particles into small, slower- or non-sinking pieces (via sloppy feeding, coprorhexy if fragmenting faeces),[52] retarding POC export. This releases dissolved organic carbon (DOC) either directly from cells or indirectly via bacterial solubilisation (yellow circle around DOC). Bacteria can then remineralise the DOC to DIC (CO2, microbial gardening).[51]
The biological carbon pump is one of the chief determinants of the vertical distribution of carbon in the oceans and therefore of the surface partial pressure of CO2 governing air-sea CO2 exchange.[53] It comprises phytoplankton cells, their consumers and the bacteria that assimilate their waste and plays a central role in the global carbon cycle by delivering carbon from the atmosphere to the deep sea, where it is concentrated and sequestered for centuries.[54] Photosynthesis by phytoplankton lowers the partial pressure of CO2 in the upper ocean, thereby facilitating the absorption of CO2 from the atmosphere by generating a steeper CO2 gradient.[55] It also results in the formation of particulate organic carbon (POC) in the euphotic layer of the epipelagic zone (0–200 m depth). The POC is processed by microbes, zooplankton and their consumers into fecal pellets, organic aggregates ("marine snow") and other forms, which are thereafter exported to the mesopelagic (200–1000 m depth) and bathypelagic zones by sinking and vertical migration by zooplankton and fish.[56] Although primary production includes both dissolved and particulate organic carbon (DOC and POC respectively), only POC leads to efficient carbon export to the ocean interior, whereas the DOC fraction in surface waters is mostly recycled by bacteria.[57] However, a more biologically resistant DOC fraction produced in the euphotic zone (accounting for 15–20% of net community productivity), is not immediately mineralized by microbes and accumulates in the ocean surface as biologically semi-labile DOC.[58] This semi-labile DOC undergoes net export to the deep ocean, thus constituting a dynamic part of the biological carbon pump.[59] The efficiency of DOC production and export varies across oceanographic regions, being more prominent in the oligotrophic subtropical oceans.[60] The overall efficiency of the biological carbon pump is mostly controlled by the export of POC.[57][61]
Marine snow
[edit]
Most carbon incorporated in organic and inorganic biological matter is formed at the sea surface where it can then start sinking to the ocean floor. The deep ocean gets most of its nutrients from the higher water column when they sink down in the form of marine snow. This is made up of dead or dying animals and microbes, fecal matter, sand and other inorganic material.[63] A single phytoplankton cell has a sinking rate around one metre per day. Given that the average depth of the ocean is about four kilometres, it can take over ten years for these cells to reach the ocean floor. However, through processes such as coagulation and expulsion in predator fecal pellets, these cells form aggregates. These aggregates, known as marine snow, have sinking rates orders of magnitude greater than individual cells and complete their journey to the deep in a matter of days.[8]
In the diagram on the right, phytoplankton fix CO2 in the euphotic zone using solar energy and produce particulate organic carbon (POC). POC formed in the euphotic zone is processed by microbes, zooplankton and their consumers into organic aggregates (marine snow), which is thereafter exported to the mesopelagic (200–1000 m depth) and bathypelagic zones by sinking and vertical migration by zooplankton and fish. Export flux is defined as the sedimentation out of the surface layer (at approximately 100 m depth) and sequestration flux is the sedimentation out of the mesopelagic zone (at approximately 1000 m depth). A portion of the POC is respired back to CO2 in the oceanic water column at depth, mostly by heterotrophic microbes and zooplankton, thus maintaining a vertical gradient in concentration of dissolved inorganic carbon (DIC). This deep-ocean DIC returns to the atmosphere on millennial timescales through thermohaline circulation. Between 1% and 40% of the primary production is exported out of the euphotic zone, which attenuates exponentially towards the base of the mesopelagic zone and only about 1% of the surface production reaches the sea floor.[61][64][56]
Of the 50–60 Pg of carbon fixed annually, roughly 10% leaves the surface mixed layer of the oceans, while less than 0.5% of eventually reaches the sea floor.[8] Most is retained in regenerated production in the euphotic zone and a significant portion is remineralized in midwater processes during particle sinking. The portion of carbon that leaves the surface mixed layer of the ocean is sometimes considered "sequestered", and essentially removed from contact with the atmosphere for many centuries.[64] However, work also finds that, in regions such as the Southern Ocean, much of this carbon can quickly (within decades) come back into contact with the atmosphere.[65]
Budget calculations of the biological carbon pump are based on the ratio between sedimentation (carbon export) and remineralization (release to the atmosphere).[62] It has been estimated that sinking particles export up to 25% of the carbon captured by phytoplankton in the surface ocean to deeper water layers.[66] About 20% of this export (5% of surface values) is buried in the ocean sediments [67] mainly due to their mineral ballast.[68] During the sinking process, these organic particles are hotspots of microbial activity and represent important loci for organic matter mineralization and nutrient redistribution in the water column.[69][70][62]
Biomineralization
[edit]Ballast minerals
[edit]| Part of a series related to |
| Biomineralization |
|---|
 |
Observations have shown that fluxes of ballast minerals (calcium carbonate, opal, and lithogenic material) and organic carbon fluxes are closely correlated in the bathypelagic zones of the ocean.[68] A large fraction of particulate organic matter occurs in the form of marine snow aggregates (>0.5 mm) composed of phytoplankton, detritus, inorganic mineral grains, and fecal pellets in the ocean.[71] Formation and sinking of these aggregates drive the biological carbon pump via export and sedimentation of organic matter from the surface mixed layer to the deep ocean and sediments. The fraction of organic matter that leaves the upper mixed layer of the ocean is, among other factors, determined by the sinking velocity and microbial remineralisation rate of these aggregates. Recent observations have shown that the fluxes of ballast minerals (calcium carbonate, opal, and lithogenic material) and the organic carbon fluxes are closely correlated in the bathypelagic zones of the ocean. This has led to the hypothesis that organic carbon export is determined by the presence of ballast minerals within settling aggregates.[72][73][74][68]
Mineral ballasting is associated with about 60% of the flux of particulate organic carbon (POC) in the high-latitude North Atlantic, and with about 40% of the flux in the Southern Ocean.[75] Strong correlations exist also in the deep ocean between the presence of ballast minerals and the flux of POC. This suggests ballast minerals enhance POC flux by increasing the sink rate of ballasted aggregates. Ballast minerals could additionally provide aggregated organic matter some protection from degradation.[76]
It has been proposed that organic carbon is better preserved in sinking particles due to increased aggregate density and sinking velocity when ballast minerals are present and/or via protection of the organic matter due to quantitative association to ballast minerals.[72][73][74] In 2002, Klaas and Archer observed that about 83% of the global particulate organic carbon (POC) fluxes were associated with carbonate, and suggested carbonate was a more efficient ballast mineral as compared to opal and terrigenous material. They hypothesized that the higher density of calcium carbonate compared to that of opal and the higher abundance of calcium carbonate relative to terrigenous material might be the reason for the efficient ballasting by calcium carbonate. However, the direct effects of ballast minerals on sinking velocity and degradation rates in sinking aggregates are still unclear.[74][68]
A 2008 study demonstrated copepod fecal pellets produced on a diet of diatoms or coccolithophorids show higher sinking velocities as compared to pellets produced on a nanoflagellate diet.[77] Carbon-specific respiration rates in pellets, however, were similar and independent of mineral content. These results suggest differences in mineral composition do not lead to differential protection of POC against microbial degradation, but the enhanced sinking velocities may result in up to 10-fold higher carbon preservation in pellets containing biogenic minerals as compared to that of pellets without biogenic minerals[77][68]
Minerals seem to enhance the flocculation of phytoplankton aggregates [78][79] and may even act as a catalyst in aggregate formation.[80] However, it has also been shown that incorporation of minerals can cause aggregates to fragment into smaller and denser aggregates.[81] This can potentially lower the sinking velocity of the aggregated organic material due to the reduced aggregate sizes, and, thus, lower the total export of organic matter. Conversely, if the incorporation of minerals increases the aggregate density, its size-specific sinking velocity may also increase, which could potentially increase the carbon export. Therefore, there is still a need for better quantitative investigations of how the interactions between minerals and organic aggregates affect the degradation and sinking velocity of the aggregates and, hence, carbon sequestration in the ocean.[81][68]
Remineralisation
[edit]Remineralisation refers to the breakdown or transformation of organic matter (those molecules derived from a biological source) into its simplest inorganic forms. These transformations form a crucial link within ecosystems as they are responsible for liberating the energy stored in organic molecules and recycling matter within the system to be reused as nutrients by other organisms.[6] What fraction does escape remineralisation varies depending on the location. For example, in the North Sea, values of carbon deposition are ~1% of primary production[82] while that value is <0.5% in the open oceans on average.[83] Therefore, most of nutrients remain in the water column, recycled by the biota. Heterotrophic organisms will utilize the materials produced by the autotrophic (and chemotrophic) organisms and via respiration will remineralise the compounds from the organic form back to inorganic, making them available for primary producers again.
For most areas of the ocean, the highest rates of carbon remineralisation occur at depths between 100–1,200 m (330–3,940 ft) in the water column, decreasing down to about 1,200 m (3,900 ft) where remineralisation rates remain pretty constant at 0.1 μmol kg−1 yr−1.[84] This provides the most nutrients available for primary producers within the photic zone, though it leaves the upper surface waters starved of inorganic nutrients.[85] Most remineralisation is done with dissolved organic carbon (DOC). Studies have shown that it is larger sinking particles that transport matter down to the sea floor[86] while suspended particles and dissolved organics are mostly consumed by remineralisation.[87] This happens in part due to the fact that organisms must typically ingest nutrients smaller than they are, often by orders of magnitude.[88] With the microbial community making up 90% of marine biomass,[89] it is particles smaller than the microbes (on the order of 10−6) that will be taken up for remineralisation.[90]
Key role of phytoplankton
[edit]

| Part of a series on |
| Biogeochemical cycles |
|---|
 |
Marine phytoplankton perform half of all photosynthesis on Earth [91] and directly influence global biogeochemical cycles and the climate, yet how they will respond to future global change is unknown. Carbon dioxide is one of the principal drivers of global change and has been identified as one of the major challenges in the 21st century.[92] Carbon dioxide (CO2) generated during anthropogenic activities such as deforestation and burning of fossil fuels for energy generation rapidly dissolves in the surface ocean and lowers seawater pH, while CO2 remaining in the atmosphere increases global temperatures and leads to increased ocean thermal stratification. While CO2 concentration in the atmosphere is estimated to be about 270 ppm before the industrial revolution, it has currently increased to about 400 ppm [93] and is expected to reach 800–1000 ppm by the end of this century according to the "business as usual" CO2 emission scenario.[94][61]
Marine ecosystems are a major sink for atmospheric CO2 and take up similar amount of CO2 as terrestrial ecosystems, currently accounting for the removal of nearly one third of anthropogenic CO2 emissions from the atmosphere.[93][94] The net transfer of CO2 from the atmosphere to the oceans and then sediments, is mainly a direct consequence of the combined effect of the solubility and the biological pump.[95] While the solubility pump serves to concentrate dissolved inorganic carbon (CO2 plus bicarbonate and carbonate ions) in the deep oceans, the biological carbon pump (a key natural process and a major component of the global carbon cycle that regulates atmospheric CO2 levels) transfers both organic and inorganic carbon fixed by primary producers (phytoplankton) in the euphotic zone to the ocean interior and subsequently to the underlying sediments.[95][54] Thus, the biological pump takes carbon out of contact with the atmosphere for several thousand years or longer and maintains atmospheric CO2 at significantly lower levels than would be the case if it did not exist.[96] An ocean without a biological pump, which transfers roughly 11 Gt C yr−1 into the ocean's interior, would result in atmospheric CO2 levels ~400 ppm higher than present day.[97][98][61]
Passow and Carlson defined sedimentation out of the surface layer (at approximately 100 m depth) as the "export flux" and that out of the mesopelagic zone (at approximately 1000 m depth) as the "sequestration flux".[64] Once carbon is transported below the mesopelagic zone, it remains in the deep sea for 100 years or longer, hence the term "sequestration" flux. According to the modelling results of Buesseler and Boyd between 1% and 40% of the primary production is exported out of the euphotic zone,[99] which attenuates exponentially towards the base of the mesopelagic zone and only about 1% of the surface production reaches the sea floor.[100] The export efficiency of particulate organic carbon (POC) shows regional variability. For instance, in the North Atlantic, over 40% of net primary production is exported out of the euphotic zone as compared to only 10% in the South Pacific,[99] and this is driven in part by the composition of the phytoplankton community including cell size and composition (see below). Exported organic carbon is remineralized, that is, respired back to CO2 in the oceanic water column at depth, mainly by heterotrophic microbes and zooplankton. Thus, the biological carbon pump maintains a vertical gradient in the concentration of dissolved inorganic carbon (DIC), with higher values at increased ocean depth.[101] This deep-ocean DIC returns to the atmosphere on millennial timescales through thermohaline circulation.[10][61]
In 2001, Hugh et al. expressed the efficiency of the biological pump as the amount of carbon exported from the surface layer (export production) divided by the total amount produced by photosynthesis (overall production).[10] Modelling studies by Buesseler and Boyd revealed that the overall transfer efficiency of the biological pump is determined by a combination of factors: seasonality;[99] the composition of phytoplankton species; the fragmentation of particles by zooplankton; and the solubilization of particles by microbes. In addition, the efficiency of the biological pump is also dependent on the aggregation and disaggregation of organic-rich aggregates and interaction between POC aggregates and suspended "ballast" minerals.[102] Ballast minerals (silicate and carbonate biominerals and dust) are the major constituents of particles that leave the ocean surface via sinking. They are typically denser than seawater and most organic matter, thus, providing a large part of the density differential needed for sinking of the particles.[72] Aggregation of particles increases vertical flux by transforming small suspended particles into larger, rapidly-sinking ones. It plays an important role in the sedimentation of phytodetritus from surface layer phytoplankton blooms.[56] As illustrated by Turner in 2015, the vertical flux of sinking particles is mainly due to a combination of fecal pellets, marine snow and direct sedimentation of phytoplankton blooms, which are typically composed of diatoms, coccolithophorids, dinoflagellates and other plankton.[56] Marine snow comprises macroscopic organic aggregates >500 μm in size and originates from clumps of aggregated phytoplankton (phytodetritus), discarded appendicularian houses, fecal matter and other miscellaneous detrital particles,[56] Appendicularians secrete mucous feeding structures or "houses" to collect food particles and discard and renew them up to 40 times a day .[103] Discarded appendicularian houses are highly abundant (thousands per m3 in surface waters) and are microbial hotspots with high concentrations of bacteria, ciliates, flagellates and phytoplankton. These discarded houses are therefore among the most important sources of aggregates directly produced by zooplankton in terms of carbon cycling potential.[104][61]

The composition of the phytoplankton community in the euphotic zone largely determines the quantity and quality of organic matter that sinks to depth.[100] The main functional groups of marine phytoplankton that contribute to export production include nitrogen fixers (diazotrophic cyanobacteria), silicifiers (diatoms) and calcifiers (coccolithophores). Each of these phytoplankton groups differ in the size and composition of their cell walls and coverings, which influence their sinking velocities.[105] For example, autotrophic picoplankton (0.2–2 μm in diameter)—which include taxa such as cyanobacteria (e.g., Prochlorococcus spp. and Synechococcus spp.) and prasinophytes (various genera of eukaryotes <2 μm)—are believed to contribute much less to carbon export from surface layers due to their small size, slow sinking velocities (<0.5 m/day) and rapid turnover in the microbial loop.[105][106] In contrast, larger phytoplankton cells such as diatoms (2–500 μm in diameter) are very efficient in transporting carbon to depth by forming rapidly sinking aggregates.[64] They are unique among phytoplankton, because they require Si in the form of silicic acid (Si(OH)4) for growth and production of their frustules, which are made of biogenic silica (bSiO2) and act as ballast.[105][107] According to the reports of Miklasz and Denny,[108] the sinking velocities of diatoms can range from 0.4 to 35 m/day.[105][107][108] Analogously, coccolithophores are covered with calcium carbonate plates called 'coccoliths', which are central to aggregation and ballasting, producing sinking velocities of nearly 5 m/day.[64][105] Although it has been assumed that picophytoplankton, characterizing vast oligotrophic areas of the ocean,[100] do not contribute substantially to the particulate organic carbon (POC) flux, in 2007 Richardson and Jackson suggested that all phytoplankton, including picoplankton cells, contribute equally to POC export.[106] They proposed alternative pathways for picoplankton carbon cycling, which rely on aggregation as a mechanism for both direct sinking (the export of picoplankton as POC) and mesozooplankton- or large filter feeder-mediated sinking of picoplankton-based production.[61]
Zooplankton grazing
[edit]Sloppy feeding
[edit]
DOC = dissolved organic carbon
POC = particulate organic carbon.
Adapted from Møller et al. (2005),[109] Saba et al. (2009)[110] and Steinberg et al. (2017).[111]


Collected from marine snow catchers (a–c) and sediment traps (d–f).
Morphological classes: (a, d) round, (b, e) cylindrical, and (c, f) ovoid.
Scale bar = 0.5 mm
In addition to linking primary producers to higher trophic levels in marine food webs, zooplankton also play an important role as "recyclers" of carbon and other nutrients that significantly impact marine biogeochemical cycles, including the biological pump. This is particularly the case with copepods and krill, and is especially important in oligotrophic waters of the open ocean. Through sloppy feeding, excretion, egestion, and leaching of fecal pellets, zooplankton release dissolved organic matter (DOM) which controls DOM cycling and supports the microbial loop. Absorption efficiency, respiration, and prey size all further complicate how zooplankton are able to transform and deliver carbon to the deep ocean.[111]
Excretion and sloppy feeding (the physical breakdown of food source) make up 80% and 20% of crustacean zooplankton-mediated DOM release respectively.[113] In the same study, fecal pellet leaching was found to be an insignificant contributor. For protozoan grazers, DOM is released primarily through excretion and egestion and gelatinous zooplankton can also release DOM through the production of mucus. Leaching of fecal pellets can extend from hours to days after initial egestion and its effects can vary depending on food concentration and quality.[114][115] Various factors can affect how much DOM is released from zooplankton individuals or populations.
Fecal pellets
[edit]The fecal pellets of zooplankton can be important vehicles for the transfer of particulate organic carbon (POC) to the deep ocean, often making large contributions to the carbon sequestration. The size distribution of the copepod community indicates high numbers of small fecal pellets are produced in the epipelagic. However, small fecal pellets are rare in the deeper layers, suggesting they are not transferred efficiently to depth. This means small fecal pellets make only minor contributions to fecal pellet fluxes in the meso- and bathypelagic, particularly in terms of carbon. In a study is focussed on the Scotia Sea, which contains some of the most productive regions in the Southern Ocean, the dominant fecal pellets in the upper mesopelagic were cylindrical and elliptical, while ovoid fecal pellets were dominant in the bathypelagic. The change in fecal pellet morphology, as well as size distribution, points to the repacking of surface fecal pellets in the mesopelagic and in situ production in the lower meso- and bathypelagic, which may be augmented by inputs of fecal pellets via zooplankton vertical migrations. This suggests the flux of carbon to the deeper layers within the Southern Ocean is strongly modulated by meso- and bathypelagic zooplankton, meaning that the community structure in these zones has a major impact on the efficiency of the fecal pellet transfer to ocean depths.[112]
Absorption efficiency (AE) is the proportion of food absorbed by plankton that determines how available the consumed organic materials are in meeting the required physiological demands.[111] Depending on the feeding rate and prey composition, variations in AE may lead to variations in fecal pellet production, and thus regulates how much organic material is recycled back to the marine environment. Low feeding rates typically lead to high AE and small, dense pellets, while high feeding rates typically lead to low AE and larger pellets with more organic content. Another contributing factor to DOM release is respiration rate. Physical factors such as oxygen availability, pH, and light conditions may affect overall oxygen consumption and how much carbon is loss from zooplankton in the form of respired CO2. The relative sizes of zooplankton and prey also mediate how much carbon is released via sloppy feeding. Smaller prey are ingested whole, whereas larger prey may be fed on more "sloppily", that is more biomatter is released through inefficient consumption.[116][117] There is also evidence that diet composition can impact nutrient release, with carnivorous diets releasing more dissolved organic carbon (DOC) and ammonium than omnivorous diets.[114]
Microbial loop
[edit]
Bacterial lysis
[edit]The microbial loop describes a trophic pathway in the marine microbial food web where dissolved organic carbon (DOC) is returned to higher trophic levels via its incorporation into bacterial biomass, and then coupled with the classic food chain formed by phytoplankton-zooplankton-nekton. The term microbial loop was coined by Farooq Azam, Tom Fenchel et al.[118] in 1983 to include the role played by bacteria in the carbon and nutrient cycles of the marine environment. In general, dissolved organic carbon is introduced into the ocean environment from bacterial lysis, the leakage or exudation of fixed carbon from phytoplankton (e.g., mucilaginous exopolymer from diatoms), sudden cell senescence, sloppy feeding by zooplankton, the excretion of waste products by aquatic animals, or the breakdown or dissolution of organic particles from terrestrial plants and soils.[119] Bacteria in the microbial loop decompose this particulate detritus to utilize this energy-rich matter for growth. Since more than 95% of organic matter in marine ecosystems consists of polymeric, high molecular weight (HMW) compounds (e.g., protein, polysaccharides, lipids), only a small portion of total dissolved organic matter (DOM) is readily utilizable to most marine organisms at higher trophic levels. This means that dissolved organic carbon is not available directly to most marine organisms; marine bacteria introduce this organic carbon into the food web, resulting in additional energy becoming available to higher trophic levels.[120]
Viral shunt
[edit]

As much as 25% of the primary production from phytoplankton in the global oceans may be recycled within the microbial loop through viral shunting.[121] The viral shunt is a mechanism whereby marine viruses prevent microbial particulate organic matter (POM) from migrating up trophic levels by recycling them into dissolved organic matter (DOM), which can be readily taken up by microorganisms. The DOM recycled by the viral shunt pathway is comparable to the amount generated by the other main sources of marine DOM.[122] Viruses can easily infect microorganisms in the microbial loop due to their relative abundance compared to microbes.[123][124] Prokaryotic and eukaryotic mortality contribute to carbon nutrient recycling through cell lysis. There is evidence as well of nitrogen (specifically ammonium) regeneration. This nutrient recycling helps stimulates microbial growth.[125]
Role of mixoplankton
[edit]
Macroorganisms
[edit]Jelly fall
[edit]
Jelly-falls are marine carbon cycling events whereby gelatinous zooplankton, primarily cnidarians, sink to the seafloor and enhance carbon and nitrogen fluxes via rapidly sinking particulate organic matter.[128] These events provide nutrition to benthic megafauna and bacteria.[129][130] Jelly-falls have been implicated as a major "gelatinous pathway" for the sequestration of labile biogenic carbon through the biological pump.[131] These events are common in protected areas with high levels of primary production and water quality suitable to support cnidarian species. These areas include estuaries and several studies have been conducted in fjords of Norway.[130]

Whale pump
[edit]
Whales and other marine mammals also enhance primary productivity in their feeding areas by concentrating nitrogen near the surface through the release of flocculent fecal plumes.[133][134] For example, whales and seals may be responsible for replenishing more nitrogen in the Gulf of Maine's euphotic zone than the input of all rivers combined. This upward whale pump played a much larger role before industrial fishing devastated marine mammal stocks, when recycling of nitrogen was likely more than three times the atmospheric nitrogen input.[133]
The biological pump mediates the removal of carbon and nitrogen from the euphotic zone through the downward flux of aggregates, feces, and vertical migration of invertebrates and fish.[135] Copepods and other zooplankton produce sinking fecal pellets and contribute to downward transport of dissolved and particulate organic matter by respiring and excreting at depth during migration cycles, thus playing an important role in the export of nutrients (N, P, and Fe) from surface waters.[136][137][133]
Zooplankton feed in the euphotic zone and export nutrients via sinking fecal pellets, and vertical migration. Fish typically release nutrients at the same depth at which they feed. Excretion for marine mammals, tethered to the surface for respiration, is expected to be shallower in the water column than where they feed.[133]
Marine mammals provide important ecosystem services. On a global scale, they can influence climate, through fertilization events and the export of carbon from surface waters to the deep sea through sinking whale carcasses.[138] In coastal areas, whales retain nutrients locally, increasing ecosystem productivity and perhaps raising the carrying capacity for other marine consumers, including commercial fish species.[133] It has been estimated that, in terms of carbon sequestration, one whale is equivalent to thousands of trees.[139]
Vertical migrations
[edit]Diel vertically migrating krill, salps, smaller zooplankton and fish can actively transport carbon to depth by consuming POC in the surface layer at night, and metabolising it at their daytime, mesopelagic residence depths. Depending on species life history, active transport may occur on a seasonal basis as well.[51]
Without vertical migration the biological pump wouldn't be nearly as efficient. Organisms migrate up to feed at night so when they migrate back to depth during the day they defecate large sinking fecal pellets. Whilst some larger fecal pellets can sink quite fast, the speed that organisms move back to depth is still faster. At night organisms are in the top 100 metres of the water column, but during the day they move down to between 800 and 1000 metres. If organisms were to defecate at the surface it would take the fecal pellets days to reach the depth that they reach in a matter of hours. Therefore, by releasing fecal pellets at depth they have almost 1000 metres less to travel to get to the deep ocean. This is something known as active transport. The organisms are playing a more active role in moving organic matter down to depths. Because a large majority of the deep sea, especially marine microbes, depends on nutrients falling down, the quicker they can reach the ocean floor the better.[63]
Zooplankton and salps play a large role in the active transport of fecal pellets. 15–50% of zooplankton biomass is estimated to migrate, accounting for the transport of 5–45% of particulate organic nitrogen to depth.[63] Salps are large gelatinous plankton that can vertically migrate 800 metres and eat large amounts of food at the surface. They have a very long gut retention time, so fecal pellets usually are released at maximum depth. Salps are also known for having some of the largest fecal pellets. Because of this they have a very fast sinking rate, small detritus particles are known to aggregate on them. This makes them sink that much faster. So while currently there is still much research being done on why organisms vertically migrate, it is clear that vertical migration plays a large role in the active transport of dissolved organic matter to depth.[140]
Lipid pump
[edit]The lipid pump sequesters carbon from the ocean's surface to deeper waters via lipids associated with overwintering vertically migratory zooplankton. Lipids are a class of hydrocarbon rich, nitrogen and phosphorus deficient compounds essential for cellular structures. The lipid associated carbon enters the deep ocean as carbon dioxide produced by respiration of lipid reserves and as organic matter from the mortality of zooplankton. Compared to the more general biological pump, the lipid pump also results in a lipid shunt, where other nutrients like nitrogen and phosphorus that are consumed in excess must be excreted back to the surface environment, and thus are not removed from the surface mixed layer of the ocean.[141] This means that the carbon transported by the lipid pump does not limit the availability of essential nutrients in the ocean surface. Carbon sequestration via the lipid pump is therefore decoupled from nutrient removal, allowing carbon uptake by oceanic primary production to continue. In the Biological Pump, nutrient removal is always coupled to carbon sequestration; primary production is limited as carbon and nutrients are transported to depth together in the form of organic matter.[141] The contribution of the lipid pump to the sequestering of carbon in the deeper waters of the ocean can be substantial: the carbon transported below 1,000 metres (3,300 ft) by copepods of the genus Calanus in the Arctic Ocean almost equals that transported below the same depth annually by particulate organic carbon (POC) in this region.[142] A significant fraction of this transported carbon would not return to the surface due to respiration and mortality. Research is ongoing to more precisely estimate the amount that remains at depth.[141][142][143] The export rate of the lipid pump may vary from 1–9.3 g C m−2 y−1 across temperate and subpolar regions containing seasonally-migrating zooplankton.[143] The role of zooplankton, and particularly copepods, in the food web is crucial to the survival of higher trophic level organisms whose primary source of nutrition is copepods. With warming oceans and increasing melting of ice caps due to climate change, the organisms associated with the lipid pump may be affected, thus influencing the survival of many commercially important fish and endangered marine mammals.[144][145][146] As a new and previously unquantified component of oceanic carbon sequestration, further research on the lipid pump can improve the accuracy and overall understanding of carbon fluxes in global oceanic systems.[141][142][143]
Bioluminescent shunt
[edit]
Luminous bacteria in light organ symbioses are successively acquired by host (squid, fish) from the seawater while they are juveniles, then regularly released into the ocean.[147]
In the diagram on the right, depending on the light organ position, luminous bacteria are released from their guts into fecal pellets or directly into the seawater (step 1). Motile luminous bacteria colonize organic matter sinking along the water column. Bioluminescent bacteria colonising fecal pellets and particles influence zooplankton consumption rates. Such visual markers increase detection ("bait hypothesis"), attraction and finally predation by upper trophic levels (step 2). In the mesopelagic, zooplankton and their predators feed on sinking luminous particles and fecal pellets, which form either aggregates (repackaging) of faster sinking rates or fragment organic matter (due to sloppy feeding) with slower sinking rates (step 3).[147]
Filter feeders also aggregate sinking organic matter without particular visual detection and selection of luminous matter. Diel (and seasonal) vertical migrators feeding on luminous food metabolize and release glowing fecal pellets from the surface to the mesopelagic zone (step 4). This implies bioluminescent bacteria dispersion at large spatial scales, for zooplankton or even some fish actively swimming long distances. Luminous bacteria attached to particles sink down to the seafloor, and sediment can be resuspended by oceanographic physical conditions (step 5) and consumed by epi-benthic organisms. Instruments are (a) plankton net, (b) fish net, (c) Niskin water sampler, (d) bathyphotometer, (e) sediment traps, (f) autonomous underwater vehicles, (g) photomultiplier module, (h) astrophysics optical modules ANTARES and (i–j) remotely operated vehicles.[147]
Quantification
[edit]
The geologic component of the carbon cycle operates slowly in comparison to the other parts of the global carbon cycle. It is one of the most important determinants of the amount of carbon in the atmosphere, and thus of global temperatures.[149]
As the biological pump plays an important role in the Earth's carbon cycle, significant effort is spent quantifying its strength. However, because they occur as a result of poorly constrained ecological interactions usually at depth, the processes that form the biological pump are difficult to measure. A common method is to estimate primary production fuelled by nitrate and ammonium as these nutrients have different sources that are related to the remineralisation of sinking material. From these it is possible to derive the so-called f-ratio, a proxy for the local strength of the biological pump. Applying the results of local studies to the global scale is complicated by the role the ocean's circulation plays in different ocean regions.[150]
Effects of climate change
[edit]
Changes in land use, the combustion of fossil fuels, and the production of cement have led to an increase in CO2 concentration in the atmosphere. At present, about one third (approximately 2 Pg C y−1 = 2 × 1015 grams of carbon per year)[151][152][unreliable source?] of anthropogenic emissions of CO2 may be entering the ocean, but this is quite uncertain.[153] Some research suggests that a link between elevated CO2 and marine primary production exists.[154]

.png)
Climate change may affect the biological pump in the future by warming and stratifying the surface ocean. It is believed that this could decrease the supply of nutrients to the euphotic zone, reducing primary production there. Also, changes in the ecological success of calcifying organisms caused by ocean acidification may affect the biological pump by altering the strength of the hard tissues pump.[157] This may then have a "knock-on" effect on the soft tissues pump because calcium carbonate acts to ballast sinking organic material.[158]
The second diagram on the right shows some possible effects of sea ice decline and permafrost thaw on Arctic carbon fluxes. On land, plants take up carbon while microorganisms in the soil produce methane and respire CO2. Lakes are net emitters of methane, and organic and inorganic carbon (dissolved and particulate) flow into the ocean through freshwater systems. In the ocean, methane can be released from thawing subsea permafrost, and CO2 is absorbed due to an undersaturation of CO2 in the water compared with the atmosphere. In addition, multiple fluxes are closely associated to sea ice. Current best estimates of atmospheric fluxes are given in Tg C year−1, where available. Note that the emission estimate for lakes is for the area North of ~50º N rather than the narrower definition of arctic tundra for the other terrestrial fluxes. When available, uncertainty ranges are shown in brackets. The arrows do not represent the size of each flux.[156][155]
The biological pump is thought to have played significant roles in atmospheric CO2 fluctuations during past glacial-interglacial periods. However, it is not yet clear how the biological pump will respond to future climate change.[56] For such predictions to be reasonable, it is important to first decipher the response of phytoplankton, one of the key components of the biological pump to future changes in atmospheric CO2. Due to their phylogenetic diversity, different phytoplankton taxa will likely respond to climate change in different ways.[105] For instance, a decrease in the abundance of diatom is expected due to increased stratification in the future ocean.[159] Diatoms are highly efficient in transporting carbon to depths by forming large, rapidly sinking aggregates and their reduced numbers could in turn lead to decreased carbon export.[64]
Further, decreased ocean pH due to ocean acidification may thwart the ability of coccolithophores to generate calcareous plates, potentially affecting the biological pump;[105] however, it appears that some species are more sensitive than others.[160] Thus, future changes in the relative abundance of these or other phytoplankton taxa could have a marked impact on total ocean productivity, subsequently affecting ocean biogeochemistry and carbon storage.
A 2015 study determined that coccolithophore concentrations in the North Atlantic have increased by an order of magnitude since the 1960s and an increase in absorbed CO2, as well as temperature, were modeled to be the most likely cause of this increase.[161]
In a 2017 study, scientists used species distribution modelling (SDM) to predict the future global distribution of two phytoplankton species important to the biological pump: the diatom Chaetoceros diadema and the coccolithophore Emiliania huxleyi.[162] They employed environmental data described in the IPCC Representative Concentration Pathways scenario 8.5, which predicts radiative forcing in the year 2100 relative to pre-industrial values. Their modelling results predicted that the total ocean area covered by C. diadema and E. huxleyi would decline by 8% and 16%, respectively, under the examined climate scenario. They predicted changes in the range and distribution of these two phytoplankton species under these future ocean conditions, if realized, might result in reduced contribution to carbon sequestration via the biological pump.[61] In 2019, a study indicated that at current rates of seawater acidification, we could see Antarctic phytoplanktons smaller and less effective at storing carbon before the end of the century.[163]
Monitoring
[edit]
Monitoring the biological pump is critical to understanding how the Earth's carbon cycle is changing. A variety of techniques are used to monitor the biological pump, which can be deployed from various platforms such as ships, autonomous vehicles, and satellites. At present, satellite remote sensing is the only tool available for viewing the entire surface ocean at high temporal and spatial scales.[4]
Needed research
[edit]
to properly understand the biological pump [166]
Multidisciplinary observations are still needed in the deep water column to properly understand the biological pump:[166]
- Physics: stratification affects particle sinking; understanding the origin of the particles and the residence time of the DIC from particle remineralization in the deep ocean requires measurement of advection and mixing.[166]
- Biogeochemistry: export/mixing down of particulate and dissolved organic matter from the surface layer determines labile organic matter arriving at the seafloor, which is either respired by seafloor biota or stored for longer times in the sediment.[166]
- Biology and ecosystems: zooplankton and microorganisms break down and remineralize sinking particles in the water column. Exported organic matter feeds all water column and benthic biota (zooplankton, benthic invertebrates, microbes) sustaining their biomass, density, and biodiversity.[166]
See also
[edit]References
[edit]- ^ a b Sigman DM & GH Haug. 2006. The biological pump in the past. In: Treatise on Geochemistry; vol. 6, (ed.). Pergamon Press, pp. 491-528
- ^ a b Hain, M.P.; Sigman, D.M.; Haug, G.H. (2014). "The Biological Pump in the Past". Treatise on Geochemistry (PDF). Vol. 8 (2 ed.). pp. 485–517. doi:10.1016/B978-0-08-095975-7.00618-5. ISBN 978-0-08-098300-4. Retrieved 1 June 2015.
- ^ Nowicki, Michael; DeVries, Tim; Siegel, David A. (March 2022). "Quantifying the Carbon Export and Sequestration Pathways of the Ocean's Biological Carbon Pump". Global Biogeochemical Cycles. 36 (3). Bibcode:2022GBioC..3607083N. doi:10.1029/2021GB007083. S2CID 246458736.
- ^ a b c d e f g h Brewin, Robert J.W.; Sathyendranath, Shubha; Platt, Trevor; Bouman, Heather; et al. (2021). "Sensing the ocean biological carbon pump from space: A review of capabilities, concepts, research gaps and future developments". Earth-Science Reviews. 217 103604. Elsevier BV. Bibcode:2021ESRv..21703604B. doi:10.1016/j.earscirev.2021.103604. hdl:10871/125469. ISSN 0012-8252. S2CID 233682755.
 Modified material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
Modified material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- ^ Volk, Tyler; Hoffert, Martin I. (2013). "Ocean Carbon Pumps: Analysis of Relative Strengths and Efficiencies in Ocean-Driven Atmospheric CO2 Changes". The Carbon Cycle and Atmospheric CO2: Natural Variations Archean to Present. Geophysical Monograph Series. pp. 99–110. doi:10.1029/GM032p0099. ISBN 978-1-118-66432-2.
- ^ a b Sarmiento, Jorge L. (17 July 2013). Ocean Biogeochemical Dynamics. Princeton University Press. p. 526. ISBN 978-1-4008-4907-9.
- ^ Middelburg, Jack J. (2019). "The Return from Organic to Inorganic Carbon". Marine Carbon Biogeochemistry. SpringerBriefs in Earth System Sciences. pp. 37–56. doi:10.1007/978-3-030-10822-9_3. ISBN 978-3-030-10821-2. S2CID 104330175. Modified text was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- ^ a b c d e f g de la Rocha, C.L.; Passow, U. (2014). "The Biological Pump". Treatise on Geochemistry. pp. 93–122. doi:10.1016/B978-0-08-095975-7.00604-5. ISBN 978-0-08-098300-4.
- ^ a b Boscolo-Galazzo, F.; Crichton, K.A.; Barker, S.; Pearson, P.N. (2018). "Temperature dependency of metabolic rates in the upper ocean: A positive feedback to global climate change?". Global and Planetary Change. 170: 201–212. Bibcode:2018GPC...170..201B. doi:10.1016/j.gloplacha.2018.08.017. Modified text was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- ^ a b c d Ducklow, Hugh; Steinberg, Deborah; Buesseler, Ken (2001). "Upper Ocean Carbon Export and the Biological Pump". Oceanography. 14 (4): 50–58. doi:10.5670/oceanog.2001.06. Modified text was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- ^ Monroy, P., Hernández-García, E., Rossi, V. and López, C. (2017) "Modeling the dynamical sinking of biogenic particles in oceanic flow". Nonlinear Processes in Geophysics, 24(2): 293–305. doi:10.5194/npg-24-293-2017. Modified text was copied from this source, which is available under a Creative Commons Attribution 3.0 International License.
- ^ Simon, M., Grossart, H., Schweitzer, B. and Ploug, H. (2002) "Microbial ecology of organic aggregates in aquatic ecosystems". Aquatic microbial ecology, 28: 175–211. doi:10.3354/ame028175.
- ^ Sigman, D.M.; Hain, M.P. (2012). "The Biological Productivity of the Ocean" (PDF). Nature Education Knowledge. 3 (6): 1–16. Retrieved 1 June 2015.
The value of NEP [Net Ecosystem Production] depends on the boundaries defined for the ecosystem. If one considers the sunlit surface ocean down to the 1% light level (the "euphotic zone") over the course of an entire year, then NEP is equivalent to the particulate organic carbon sinking into the dark ocean interior plus the dissolved organic carbon being circulated out of the euphotic zone. In this case, NEP is also often referred to as "export production" (or "new production" (Dugdale & Goering 1967), as discussed below).
- ^ Martiny, Adam C.; Pham, Chau T. A.; Primeau, Francois W.; Vrugt, Jasper A.; Moore, J. Keith; Levin, Simon A.; Lomas, Michael W. (April 2013). "Strong latitudinal patterns in the elemental ratios of marine plankton and organic matter". Nature Geoscience. 6 (4): 279–283. Bibcode:2013NatGe...6..279M. doi:10.1038/NGEO1757. S2CID 5677709.
- ^ a b c Heinrichs, Mara E.; Mori, Corinna; Dlugosch, Leon (2020). "Complex Interactions Between Aquatic Organisms and Their Chemical Environment Elucidated from Different Perspectives". YOUMARES 9 - the Oceans: Our Research, Our Future. pp. 279–297. doi:10.1007/978-3-030-20389-4_15. ISBN 978-3-030-20388-7. S2CID 210308256. Modified text was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- ^ Carlson CA (2002) [books.google.com/books?id=D6TMKZOgldAC&q=%22Production+and+removal+processes%22&pg=PA91 "Production and removal processes"]. In: Hansell DA, Carlson CA (eds) Biogeochemistry of marine dissolved organic matter. Academic Press, San Diego, pages 91–151. ISBN 9780123238412.
- ^ Capelle, David W.; Kuzyk, Zou Zou A.; Papakyriakou, Tim; Guéguen, Céline; Miller, Lisa A.; MacDonald, Robie W. (2020). "Effect of terrestrial organic matter on ocean acidification and CO2 flux in an Arctic shelf sea". Progress in Oceanography. 185 102319. Bibcode:2020PrOce.18502319C. doi:10.1016/j.pocean.2020.102319. hdl:1993/34767. Modified text was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- ^ Hedges, John I. (1992). "Global biogeochemical cycles: progress and problems". Marine Chemistry. 39 (1–3). Elsevier BV: 67–93. Bibcode:1992MarCh..39...67H. doi:10.1016/0304-4203(92)90096-s. ISSN 0304-4203.
- ^ Zeebe, Richard E.; Wolf-Gladrow, Dieter A. (2001). CO2 in seawater: equilibrium, kinetics, isotopes. Amsterdam. p. 65. ISBN 978-0-08-052922-6. OCLC 246683387.
{{cite book}}: CS1 maint: location missing publisher (link) - ^ Rost, Björn; Riebesell, Ulf (2004). "Coccolithophores and the biological pump: Responses to environmental changes". Coccolithophores. Berlin, Heidelberg: Springer Berlin Heidelberg. pp. 99–125. doi:10.1007/978-3-662-06278-4_5. ISBN 978-3-642-06016-8.
- ^ Zeebe, Richard E. (30 May 2012). "History of Seawater Carbonate Chemistry, Atmospheric CO2, and Ocean Acidification". Annual Review of Earth and Planetary Sciences. 40 (1). Annual Reviews: 141–165. Bibcode:2012AREPS..40..141Z. doi:10.1146/annurev-earth-042711-105521. ISSN 0084-6597.
- ^ a b Hansell, Dennis A.; Carlson, Craig A. (12 August 2013). "Localized refractory dissolved organic carbon sinks in the deep ocean". Global Biogeochemical Cycles. 27 (3). American Geophysical Union (AGU): 705–710. doi:10.1002/gbc.20067. ISSN 0886-6236. S2CID 17175370.
- ^ a b Hansell, Dennis A. (3 January 2013). "Recalcitrant Dissolved Organic Carbon Fractions". Annual Review of Marine Science. 5 (1). Annual Reviews: 421–445. doi:10.1146/annurev-marine-120710-100757. ISSN 1941-1405. PMID 22881353.
- ^ Williams, Peter M.; Druffel, Ellen R. M. (1987). "Radiocarbon in dissolved organic matter in the central North Pacific Ocean". Nature. 330 (6145). Springer Science and Business Media LLC: 246–248. Bibcode:1987Natur.330..246W. doi:10.1038/330246a0. ISSN 0028-0836. S2CID 4329024.
- ^ Druffel, E. R. M.; Griffin, S.; Coppola, A. I.; Walker, B. D. (28 May 2016). "Radiocarbon in dissolved organic carbon of the Atlantic Ocean". Geophysical Research Letters. 43 (10). American Geophysical Union (AGU): 5279–5286. Bibcode:2016GeoRL..43.5279D. doi:10.1002/2016gl068746. ISSN 0094-8276. S2CID 56069589.
- ^ Stramska, Malgorzata; Cieszyńska, Agata (18 July 2015). "Ocean colour estimates of particulate organic carbon reservoirs in the global ocean – revisited". International Journal of Remote Sensing. 36 (14). Informa UK Limited: 3675–3700. Bibcode:2015IJRS...36.3675S. doi:10.1080/01431161.2015.1049380. ISSN 0143-1161. S2CID 128524215.
- ^ Wickland, D.E., Plummer, S. and Nakajima, M. (October 2013). "CEOS strategy for carbon observations from space". In: International Conference Towards a Global Carbon Observing System: Progresses and Challenges, 1:. 2.
- ^ Sarmiento, Jorge L. (1 January 2006). Ocean Biogeochemical Dynamics. Princeton University Press. doi:10.1515/9781400849079. ISBN 978-1-4008-4907-9.
- ^ Longhurst, Alan; Sathyendranath, Shubha; Platt, Trevor; Caverhill, Carla (1995). "An estimate of global primary production in the ocean from satellite radiometer data". Journal of Plankton Research. 17 (6). Oxford University Press (OUP): 1245–1271. doi:10.1093/plankt/17.6.1245. ISSN 0142-7873.
- ^ Sathyendranath, S.; Platt, T.; Brewin, Robert J.W.; Jackson, Thomas (2019). "Primary Production Distribution". Encyclopedia of Ocean Sciences. Elsevier. pp. 635–640. doi:10.1016/b978-0-12-409548-9.04304-9. ISBN 978-0-12-813082-7.
- ^ Kulk, Gemma; Platt, Trevor; Dingle, James; Jackson, Thomas; et al. (3 March 2020). "Primary Production, an Index of Climate Change in the Ocean: Satellite-Based Estimates over Two Decades". Remote Sensing. 12 (5). MDPI AG: 826. Bibcode:2020RemS...12..826K. doi:10.3390/rs12050826. hdl:11336/128979. ISSN 2072-4292.
- ^ Hopkins, Jason; Henson, Stephanie A.; Poulton, Alex J.; Balch, William M. (2019). "Regional Characteristics of the Temporal Variability in the Global Particulate Inorganic Carbon Inventory". Global Biogeochemical Cycles. 33 (11). American Geophysical Union (AGU): 1328–1338. Bibcode:2019GBioC..33.1328H. doi:10.1029/2019gb006300. ISSN 0886-6236. S2CID 210342576.
- ^ a b Feely, Richard A.; Sabine, Christopher L.; Lee, Kitack; Berelson, Will; Kleypas, Joanie; Fabry, Victoria J.; Millero, Frank J. (16 July 2004). "Impact of Anthropogenic CO 2 on the CaCO 3 System in the Oceans". Science. 305 (5682). American Association for the Advancement of Science (AAAS): 362–366. Bibcode:2004Sci...305..362F. doi:10.1126/science.1097329. ISSN 0036-8075. PMID 15256664. S2CID 31054160.
- ^ Schiebel, Ralf (24 October 2002). "Planktic foraminiferal sedimentation and the marine calcite budget". Global Biogeochemical Cycles. 16 (4). American Geophysical Union (AGU): 3–1–3–21. Bibcode:2002GBioC..16.1065S. doi:10.1029/2001gb001459. ISSN 0886-6236. S2CID 128737252.
- ^ Riebesell, Ulf; Zondervan, Ingrid; Rost, Björn; Tortell, Philippe D.; Zeebe, Richard E.; Morel, François M. M. (2000). "Reduced calcification of marine plankton in response to increased atmospheric CO2" (PDF). Nature. 407 (6802). Springer Science and Business Media LLC: 364–367. Bibcode:2000Natur.407..364R. doi:10.1038/35030078. ISSN 0028-0836. PMID 11014189. S2CID 4426501.
- ^ Mitchell, C.; Hu, C.; Bowler, B.; Drapeau, D.; Balch, W. M. (2017). "Estimating Particulate Inorganic Carbon Concentrations of the Global Ocean from Ocean Color Measurements Using a Reflectance Difference Approach". Journal of Geophysical Research: Oceans. 122 (11): 8707–8720. Bibcode:2017JGRC..122.8707M. doi:10.1002/2017JC013146.
- ^ Wilson, R. W.; Millero, F. J.; Taylor, J. R.; Walsh, P. J.; Christensen, V.; Jennings, S.; Grosell, M. (16 January 2009). "Contribution of Fish to the Marine Inorganic Carbon Cycle". Science. 323 (5912): 359–362. Bibcode:2009Sci...323..359W. doi:10.1126/science.1157972. PMID 19150840. S2CID 36321414.
- ^ Pilson MEQ. 2012. An Introduction to the Chemistry of the Sea. Cambridge University Press, pp.
- ^ Hain, M.P.; Sigman, D.M.; Haug, G.H. (2010). "Carbon dioxide effects of Antarctic stratification, North Atlantic Intermediate Water formation, and subantarctic nutrient drawdown during the last ice age: Diagnosis and synthesis in a geochemical box model". Global Biogeochemical Cycles. 24 (4): 1–19. Bibcode:2010GBioC..24.4023H. doi:10.1029/2010GB003790.
- ^ Webb, Paul (2019) Introduction to Oceanography, Chapter 12: Ocean Sediments, page 273–297, Rebus Community. Updated 2020.
- ^ H., Schlesinger, William (2013). Biogeochemistry: an analysis of global change. Bernhardt, Emily S. (3rd ed.). Waltham, Mass.: Academic Press. ISBN 978-0-12-385874-0. OCLC 827935936.
{{cite book}}: CS1 maint: multiple names: authors list (link) - ^ Falkowski, P.; Scholes, R. J.; Boyle, E.; Canadell, J.; Canfield, D.; Elser, J.; Gruber, N.; Hibbard, K.; Högberg, P. (13 October 2000). "The Global Carbon Cycle: A Test of Our Knowledge of Earth as a System". Science. 290 (5490): 291–296. Bibcode:2000Sci...290..291F. doi:10.1126/science.290.5490.291. ISSN 0036-8075. PMID 11030643.
- ^ Raven, J. A.; P. G. Falkowski (1999). "Oceanic sinks for atmospheric CO2". Plant, Cell and Environment. 22 (6): 741–755. doi:10.1046/j.1365-3040.1999.00419.x.
- ^ Raven, J. A.; Falkowski, P. G. (1999). "Oceanic sinks for atmospheric CO2". Plant, Cell and Environment. 22 (6): 741–755. doi:10.1046/j.1365-3040.1999.00419.x.
- ^ Hain, M.P.; Sigman, D.M.; Haug, G.H (2014). "The Biological Pump in the Past". Treatise on Geochemistry. 8: 485–517. doi:10.1016/B978-0-08-095975-7.00618-5. ISBN 978-0-08-098300-4.
- ^ a b Rost, Bjorn; Reibessel, Ulf (2004). Coccolithophores and the biological pump: responses to environmental changes. Berlin, Heidelberg: Springer. ISBN 978-3-642-06016-8.
- ^ Zeebe, R.E., 2016. "The calcium carbonate counter pump: Fundamentals, evolution through time, and future feedbacks". American Geophysical Union, pp.B23A-08.
- ^ a b c Tsunogai, S.; Watanabe, S.; Sato, T. (1999). "Is there a "continental shelf pump" for the absorption of atmospheric CO2". Tellus B. 51 (3): 701–712. Bibcode:1999TellB..51..701T. doi:10.1034/j.1600-0889.1999.t01-2-00010.x.
- ^ Wollast, R. (1998). Evaluation and comparison of the global carbon cycle in the coastal zone and in the open ocean, p. 213-252. In K. H. Brink and A. R. Robinson (eds.), The Global Coastal Ocean. John Wiley & Sons.
- ^ Rippeth, T. P.; Scourse, J. D.; Uehara, K.; McKeown, S. (2008). "Impact of sea-level rise over the last deglacial transition on the strength of the continental shelf CO2 pump". Geophys. Res. Lett. 35 (24): L24604. Bibcode:2008GeoRL..3524604R. doi:10.1029/2008GL035880. S2CID 1049049.
- ^ a b c Cavan, E.L., Belcher, A., Atkinson, A., Hill, S.L., Kawaguchi, S., McCormack, S., Meyer, B., Nicol, S., Ratnarajah, L., Schmidt, K. and Steinberg, D.K. (2019) "The importance of Antarctic krill in biogeochemical cycles". Nature communications, 10(1): 1–13. doi:10.1038/s41467-019-12668-7. Modified text was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- ^ Lampitt, R. S.; Noji, T.; von Bodungen, B. (1990). "What happens to zooplankton faecal pellets? Implications for material flux". Marine Biology. 104 (1): 15–23. Bibcode:1990MarBi.104...15L. doi:10.1007/BF01313152. S2CID 86523326.
- ^ Bishop, James (2009). "Autonomous Observations of the Ocean Biological Carbon Pump". Oceanography. 22 (2): 182–193. doi:10.5670/oceanog.2009.48.
- ^ a b Chisholm, Sallie W. (1995). "The iron hypothesis: Basic research meets environmental policy". Reviews of Geophysics. 33 (S2): 1277–1286. Bibcode:1995RvGeo..33S1277C. doi:10.1029/95RG00743.
- ^ Falkowski, P.; Scholes, R. J.; Boyle, E.; Canadell, J.; Canfield, D.; Elser, J.; Gruber, N.; Hibbard, K.; Högberg, P.; Linder, S.; MacKenzie, F. T.; Moore b, 3rd; Pedersen, T.; Rosenthal, Y.; Seitzinger, S.; Smetacek, V.; Steffen, W. (2000). "The Global Carbon Cycle: A Test of Our Knowledge of Earth as a System". Science. 290 (5490): 291–296. Bibcode:2000Sci...290..291F. doi:10.1126/science.290.5490.291. PMID 11030643.
{{cite journal}}: CS1 maint: numeric names: authors list (link) - ^ a b c d e f Turner, Jefferson T. (2015). "Zooplankton fecal pellets, marine snow, phytodetritus and the ocean's biological pump". Progress in Oceanography. 130: 205–248. Bibcode:2015PrOce.130..205T. doi:10.1016/j.pocean.2014.08.005.
- ^ a b Kim, Ja-Myung; Lee, Kitack; Shin, Kyungsoon; Yang, Eun Jin; Engel, Anja; Karl, David M.; Kim, Hyun-Cheol (2011). "Shifts in biogenic carbon flow from particulate to dissolved forms under high carbon dioxide and warm ocean conditions". Geophysical Research Letters. 38 (8): n/a. Bibcode:2011GeoRL..38.8612K. doi:10.1029/2011GL047346.
- ^ Hansell, Dennis; Carlson, Craig; Repeta, Daniel; Schlitzer, Reiner (2009). "Dissolved Organic Matter in the Ocean: A Controversy Stimulates New Insights". Oceanography. 22 (4): 202–211. doi:10.5670/oceanog.2009.109. hdl:1912/3183.
- ^ Carlson, Craig A.; Ducklow, Hugh W.; Michaels, Anthony F. (1994). "Annual flux of dissolved organic carbon from the euphotic zone in the northwestern Sargasso Sea". Nature. 371 (6496): 405–408. Bibcode:1994Natur.371..405C. doi:10.1038/371405a0. S2CID 4279712.
- ^ Roshan, Saeed; Devries, Timothy (2017). "Efficient dissolved organic carbon production and export in the oligotrophic ocean". Nature Communications. 8 (1): 2036. Bibcode:2017NatCo...8.2036R. doi:10.1038/s41467-017-02227-3. PMC 5725454. PMID 29230041.
- ^ a b c d e f g h i Basu, Samarpita; MacKey, Katherine (2018). "Phytoplankton as Key Mediators of the Biological Carbon Pump: Their Responses to a Changing Climate". Sustainability. 10 (3): 869. doi:10.3390/su10030869. Modified text was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- ^ a b c Ionescu, Danny; Bizic-Ionescu, Mina; Khalili, Arzhang; Malekmohammadi, Reza; Morad, Mohammad Reza; De Beer, Dirk; Grossart, Hans-Peter (2015). "A new tool for long-term studies of POM-bacteria interactions: Overcoming the century-old Bottle Effect". Scientific Reports. 5 14706. Bibcode:2015NatSR...514706I. doi:10.1038/srep14706. PMC 4592964. PMID 26435525. Modified text was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- ^ a b c Steinberg, Deborah; Sarah Goldthwait; Dennis Hansell (2002). "Zooplankton vertical migration and the active transport of dissolved organic and inorganic nitrogen in the Sargasso Sea". Deep-Sea Research Part I. 49 (8): 1445–1461. Bibcode:2002DSRI...49.1445S. CiteSeerX 10.1.1.391.7622. doi:10.1016/S0967-0637(02)00037-7. ISSN 0967-0637.
- ^ a b c d e f Passow, U.; Carlson, CA (2012). "The biological pump in a high CO2 world". Marine Ecology Progress Series. 470: 249–271. Bibcode:2012MEPS..470..249P. doi:10.3354/meps09985.
- ^ Robinson, J.; Popova, E.E.; Yool, A.; Srokosz, M.A.; Lampitt, R.S.; Blundell, J.R. (2014). "How deep is deep enough? Ocean iron fertilization and carbon sequestration in the Southern Ocean" (PDF). Geophys. Res. Lett. 41 (7): 2489–2495. Bibcode:2014GeoRL..41.2489R. doi:10.1002/2013GL058799.
- ^ Falkowski, P. G.; Barber, R. T.; Smetacek v, V. (1998). "Biogeochemical Controls and Feedbacks on Ocean Primary Production". Science. 281 (5374): 200–206. doi:10.1126/science.281.5374.200. PMID 9660741.
- ^ Yool, Andrew; Martin, Adrian P.; Fernández, Camila; Clark, Darren R. (2007). "The significance of nitrification for oceanic new production". Nature. 447 (7147): 999–1002. Bibcode:2007Natur.447..999Y. doi:10.1038/nature05885. PMID 17581584. S2CID 4416535.
- ^ a b c d e f Iversen, M. H.; Ploug, H. "Ballast minerals and the sinking carbon flux in the ocean: Carbon-specific respiration rates and sinking velocities of macroscopic organic aggregates (Marine snow)". doi:10.5194/bgd-7-3335-2010.
{{cite journal}}: Cite journal requires|journal=(help) Modified text was copied from this source, which is available under a Creative Commons Attribution 3.0 International License.
- ^ Simon, M.; Grossart, HP; Schweitzer, B.; Ploug, H. (2002). "Microbial ecology of organic aggregates in aquatic ecosystems". Aquatic Microbial Ecology. 28: 175–211. doi:10.3354/ame028175.
- ^ Azam, F.; Fenchel, T.; Field, JG; Gray, JS; Meyer-Reil, LA; Thingstad, F. (1983). "The Ecological Role of Water-Column Microbes in the Sea". Marine Ecology Progress Series. 10: 257–263. Bibcode:1983MEPS...10..257A. doi:10.3354/meps010257.
- ^ Alldredge, Alice L.; Silver, Mary W. (1988). "Characteristics, dynamics and significance of marine snow". Progress in Oceanography. 20 (1): 41–82. Bibcode:1988PrOce..20...41A. doi:10.1016/0079-6611(88)90053-5.
- ^ a b c Armstrong, Robert A.; Lee, Cindy; Hedges, John I.; Honjo, Susumu; Wakeham, Stuart G. (2001). "A new, mechanistic model for organic carbon fluxes in the ocean based on the quantitative association of POC with ballast minerals". Deep Sea Research Part II: Topical Studies in Oceanography. 49 (1–3): 219–236. Bibcode:2001DSRII..49..219A. doi:10.1016/S0967-0645(01)00101-1.
- ^ a b Francois, Roger; Honjo, Susumu; Krishfield, Richard; Manganini, Steve (2002). "Factors controlling the flux of organic carbon to the bathypelagic zone of the ocean". Global Biogeochemical Cycles. 16 (4): 34-1 – 34-20. Bibcode:2002GBioC..16.1087F. doi:10.1029/2001GB001722. S2CID 128876389.
- ^ a b c Klaas, Christine; Archer, David E. (2002). "Association of sinking organic matter with various types of mineral ballast in the deep sea: Implications for the rain ratio". Global Biogeochemical Cycles. 16 (4): 63-1 – 63-14. Bibcode:2002GBioC..16.1116K. doi:10.1029/2001GB001765.
- ^ Le Moigne, Frédéric A. C.; Pabortsava, Katsiaryna; Marcinko, Charlotte L. J.; Martin, Patrick; Sanders, Richard J. (2014). "Where is mineral ballast important for surface export of particulate organic carbon in the ocean?". Geophysical Research Letters. 41 (23): 8460–8468. Bibcode:2014GeoRL..41.8460L. doi:10.1002/2014GL061678. PMC 4459180. PMID 26074644.
- ^ Iversen, Morten H.; Robert, Maya L. (2015). "Ballasting effects of smectite on aggregate formation and export from a natural plankton community". Marine Chemistry. 175: 18–27. Bibcode:2015MarCh.175...18I. doi:10.1016/j.marchem.2015.04.009.
- ^ a b Ploug, Helle; Iversen, Morten Hvitfeldt; Koski, Marja; Buitenhuis, Erik Theodoor (2008). "Production, oxygen respiration rates, and sinking velocity of copepod fecal pellets: Direct measurements of ballasting by opal and calcite". Limnology and Oceanography. 53 (2): 469–476. Bibcode:2008LimOc..53..469P. doi:10.4319/lo.2008.53.2.0469.
- ^ Engel, Anja; Abramson, Lynn; Szlosek, Jennifer; Liu, Zhanfei; Stewart, Gillian; Hirschberg, David; Lee, Cindy (2009). "Investigating the effect of ballasting by CaCO3 in Emiliania huxleyi, II: Decomposition of particulate organic matter". Deep Sea Research Part II: Topical Studies in Oceanography. 56 (18): 1408–1419. Bibcode:2009DSRII..56.1408E. doi:10.1016/j.dsr2.2008.11.028.
- ^ Engel, Anja; Szlosek, Jennifer; Abramson, Lynn; Liu, Zhanfei; Lee, Cindy (2009). "Investigating the effect of ballasting by CaCO3 in Emiliania huxleyi: I. Formation, settling velocities and physical properties of aggregates". Deep Sea Research Part II: Topical Studies in Oceanography. 56 (18): 1396–1407. Bibcode:2009DSRII..56.1396E. doi:10.1016/j.dsr2.2008.11.027.
- ^ Lee, Cindy; Peterson, Michael L.; Wakeham, Stuart G.; Armstrong, Robert A.; Cochran, J. Kirk; Miquel, Juan Carlos; Fowler, Scott W.; Hirschberg, David; Beck, Aaron; Xue, Jianhong (2009). "Particulate organic matter and ballast fluxes measured using time-series and settling velocity sediment traps in the northwestern Mediterranean Sea". Deep Sea Research Part II: Topical Studies in Oceanography. 56 (18): 1420–1436. Bibcode:2009DSRII..56.1420L. doi:10.1016/j.dsr2.2008.11.029.
- ^ a b Passow, Uta; de la Rocha, Christina L. (2006). "Accumulation of mineral ballast on organic aggregates". Global Biogeochemical Cycles. 20 (1): n/a. Bibcode:2006GBioC..20.1013P. doi:10.1029/2005GB002579.
- ^ Thomas, Helmuth; Bozec, Yann; Elkalay, Khalid; Baar, Hein J. W. de (14 May 2004). "Enhanced Open Ocean Storage of CO2 from Shelf Sea Pumping" (PDF). Science. 304 (5673): 1005–1008. Bibcode:2004Sci...304.1005T. doi:10.1126/science.1095491. hdl:11370/e821600e-4560-49e8-aeec-18eeb17549e3. ISSN 0036-8075. PMID 15143279. S2CID 129790522.
- ^ De La Rocha, C. L. (2006). "The Biological Pump". In Holland, Heinrich D.; Turekian, Karl K. (eds.). Treatise on Geochemistry. Vol. 6. Pergamon Press. p. 625. Bibcode:2003TrGeo...6...83D. doi:10.1016/B0-08-043751-6/06107-7. ISBN 978-0-08-043751-4.
- ^ Feely, Richard A.; Sabine, Christopher L.; Schlitzer, Reiner; Bullister, John L.; Mecking, Sabine; Greeley, Dana (1 February 2004). "Oxygen Utilization and Organic Carbon Remineralisation in the Upper Water Column of the Pacific Ocean". Journal of Oceanography. 60 (1): 45–52. Bibcode:2004JOce...60...45F. doi:10.1023/B:JOCE.0000038317.01279.aa. ISSN 0916-8370. S2CID 67846685.
- ^ Administration, US Department of Commerce, National Oceanic and Atmospheric. "How far does light travel in the ocean?". oceanservice.noaa.gov. Retrieved 29 February 2016.
{{cite web}}: CS1 maint: multiple names: authors list (link) - ^ Karl, David M.; Knauer, George A.; Martin, John H. (1 March 1988). "Downward flux of particulate organic matter in the ocean: a particle decomposition paradox". Nature. 332 (6163): 438–441. Bibcode:1988Natur.332..438K. doi:10.1038/332438a0. ISSN 0028-0836. S2CID 4356597.
- ^ Lefévre, D.; Denis, M.; Lambert, C. E.; Miquel, J. -C. (1 February 1996). "Is DOC the main source of organic matter remineralization in the ocean water column?". Journal of Marine Systems. The Coastal Ocean in a Global Change Perspective. 7 (2–4): 281–291. Bibcode:1996JMS.....7..281L. doi:10.1016/0924-7963(95)00003-8.
- ^ Schulze, Ernst-Detlef; Mooney, Harold A. (6 December 2012). Biodiversity and Ecosystem Function. Springer Science & Business Media. ISBN 978-3-642-58001-7.
- ^ "International Census of Marine Microbes (ICoMM)". www.coml.org. Census of Marine Life. Archived from the original on 17 March 2016. Retrieved 29 February 2016.
- ^ "Microbe Size - Boundless Open Textbook". Boundless. Retrieved 29 February 2016.
- ^ Baumert, Helmut Z.; Petzoldt, Thomas (2008). "The role of temperature, cellular quota and nutrient concentrations for photosynthesis, growth and light–dark acclimation in phytoplankton". Limnologica. 38 (3–4): 313–326. Bibcode:2008Limng..38..313B. doi:10.1016/j.limno.2008.06.002.
- ^ Lam, Man Kee; Lee, Keat Teong; Mohamed, Abdul Rahman (2012). "Current status and challenges on microalgae-based carbon capture". International Journal of Greenhouse Gas Control. 10: 456–469. Bibcode:2012IJGGC..10..456L. doi:10.1016/j.ijggc.2012.07.010.
- ^ a b Häder, Donat-P.; Villafañe, Virginia E.; Helbling, E. Walter (2014). "Productivity of aquatic primary producers under global climate change". Photochem. Photobiol. Sci. 13 (10): 1370–1392. doi:10.1039/C3PP50418B. hdl:11336/24725. PMID 25191675. S2CID 25740029.
- ^ a b Li, Wei; Gao, Kunshan; Beardall, John (2012). "Interactive Effects of Ocean Acidification and Nitrogen-Limitation on the Diatom Phaeodactylum tricornutum". PLOS ONE. 7 (12) e51590. Bibcode:2012PLoSO...751590L. doi:10.1371/journal.pone.0051590. PMC 3517544. PMID 23236517.
- ^ a b Hülse, Dominik; Arndt, Sandra; Wilson, Jamie D.; Munhoven, Guy; Ridgwell, Andy (2017). "Understanding the causes and consequences of past marine carbon cycling variability through models" (PDF). Earth-Science Reviews. 171: 349–382. Bibcode:2017ESRv..171..349H. doi:10.1016/j.earscirev.2017.06.004. hdl:1983/7035f071-71c5-48a5-91f6-a8286e386d21.
- ^ Hutchins, David A.; Fu, Feixue (2017). "Microorganisms and ocean global change". Nature Microbiology. 2 (6): 17058. doi:10.1038/nmicrobiol.2017.58. PMID 28540925. S2CID 23357501.
- ^ Sanders, Richard; Henson, Stephanie A.; Koski, Marja; de la Rocha, Christina L.; Painter, Stuart C.; Poulton, Alex J.; Riley, Jennifer; Salihoglu, Baris; Visser, Andre; Yool, Andrew; Bellerby, Richard; Martin, Adrian P. (2014). "The Biological Carbon Pump in the North Atlantic". Progress in Oceanography. 129: 200–218. Bibcode:2014PrOce.129..200S. doi:10.1016/j.pocean.2014.05.005. hdl:11511/31027.
- ^ Boyd, Philip W. (2015). "Toward quantifying the response of the oceans' biological pump to climate change". Frontiers in Marine Science. 2. doi:10.3389/fmars.2015.00077. S2CID 16787695.
- ^ a b c Buesseler, Ken O.; Boyd, Philip W. (2009). "Shedding light on processes that control particle export and flux attenuation in the twilight zone of the open ocean". Limnology and Oceanography. 54 (4): 1210–1232. Bibcode:2009LimOc..54.1210B. doi:10.4319/lo.2009.54.4.1210.
- ^ a b c Herndl, Gerhard J.; Reinthaler, Thomas (2013). "Microbial control of the dark end of the biological pump". Nature Geoscience. 6 (9): 718–724. Bibcode:2013NatGe...6..718H. doi:10.1038/ngeo1921. PMC 3972885. PMID 24707320.
- ^ Hofmann, M.; Schellnhuber, H.-J. (2009). "Oceanic acidification affects marine carbon pump and triggers extended marine oxygen holes". Proceedings of the National Academy of Sciences. 106 (9): 3017–3022. Bibcode:2009PNAS..106.3017H. doi:10.1073/pnas.0813384106. PMC 2642667. PMID 19218455.
- ^ de la Rocha, Christina L.; Passow, Uta (2007). "Factors influencing the sinking of POC and the efficiency of the biological carbon pump" (PDF). Deep Sea Research Part II: Topical Studies in Oceanography. 54 (5–7): 639–658. Bibcode:2007DSRII..54..639D. doi:10.1016/j.dsr2.2007.01.004.
- ^ Sato, R.; Tanaka, Y.; Ishimaru, T. (2003). "Species-specific house productivity of appendicularians". Marine Ecology Progress Series. 259: 163–172. Bibcode:2003MEPS..259..163S. doi:10.3354/meps259163.
- ^ Nishibe, Yuichiro; Takahashi, Kazutaka; Ichikawa, Tadafumi; Hidaka, Kiyotaka; Kurogi, Hiroaki; Segawa, Kyohei; Saito, Hiroaki (2015). "Degradation of discarded appendicularian houses by oncaeid copepods". Limnology and Oceanography. 60 (3): 967–976. Bibcode:2015LimOc..60..967N. doi:10.1002/lno.10061.
- ^ a b c d e f g Collins, Sinéad; Rost, Björn; Rynearson, Tatiana A. (2014). "Evolutionary potential of marine phytoplankton under ocean acidification". Evolutionary Applications. 7 (1): 140–155. Bibcode:2014EvApp...7..140C. doi:10.1111/eva.12120. PMC 3894903. PMID 24454553.
- ^ a b Richardson, T. L.; Jackson, G. A. (2007). "Small Phytoplankton and Carbon Export from the Surface Ocean". Science. 315 (5813): 838–840. Bibcode:2007Sci...315..838R. doi:10.1126/science.1133471. PMID 17289995. S2CID 45563491.
- ^ a b Ragueneau, Olivier; Schultes, Sabine; Bidle, Kay; Claquin, Pascal; Moriceau, Brivaëla (2006). "Si and C interactions in the world ocean: Importance of ecological processes and implications for the role of diatoms in the biological pump". Global Biogeochemical Cycles. 20 (4): n/a. Bibcode:2006GBioC..20.4S02R. doi:10.1029/2006GB002688.
- ^ a b Miklasz, Kevin A.; Denny, Mark W. (2010). "Diatom sinkings speeds: Improved predictions and insight from a modified Stokes' law". Limnology and Oceanography. 55 (6): 2513–2525. Bibcode:2010LimOc..55.2513M. doi:10.4319/lo.2010.55.6.2513.
- ^ Møller, EF; Thor, P.; Nielsen, TG (2003). "Production of DOC by Calanus finmarchicus, C. Glacialis and C. Hyperboreus through sloppy feeding and leakage from fecal pellets". Marine Ecology Progress Series. 262: 185–191. Bibcode:2003MEPS..262..185M. doi:10.3354/meps262185.
- ^ Saba, GK; Steinberg, DK; Bronk, DA (2009). "Effects of diet on release of dissolved organic and inorganic nutrients by the copepod Acartia tonsa". Marine Ecology Progress Series. 386: 147–161. Bibcode:2009MEPS..386..147S. doi:10.3354/meps08070.
- ^ a b c Steinberg, Deborah K.; Landry, Michael R. (2017). "Zooplankton and the Ocean Carbon Cycle". Annual Review of Marine Science. 9: 413–444. Bibcode:2017ARMS....9..413S. doi:10.1146/annurev-marine-010814-015924. PMID 27814033.
- ^ a b c Belcher, Anna; Manno, Clara; Ward, Peter; Henson, Stephanie A.; Sanders, Richard; Tarling, Geraint A. (2017). "Copepod faecal pellet transfer through the meso- and bathypelagic layers in the Southern Ocean in spring". Biogeosciences. 14 (6): 1511–1525. Bibcode:2017BGeo...14.1511B. doi:10.5194/bg-14-1511-2017. Modified text was copied from this source, which is available under a Creative Commons Attribution 3.0 International License.
- ^ Saba, Grace K.; Steinberg, Deborah K.; Bronk, Deborah A. (2011). "The relative importance of sloppy feeding, excretion, and fecal pellet leaching in the release of dissolved carbon and nitrogen by Acartia tonsa copepods". Journal of Experimental Marine Biology and Ecology. 404 (1–2): 47–56. Bibcode:2011JEMBE.404...47S. doi:10.1016/j.jembe.2011.04.013.
- ^ a b Thor, P.; Dam, HG; Rogers, DR (2003). "Fate of organic carbon released from decomposing copepod fecal pellets in relation to bacterial production and ectoenzymatic activity". Aquatic Microbial Ecology. 33: 279–288. doi:10.3354/ame033279.
- ^ Hansell, Dennis A.; Carlson, Craig A. (2 October 2014). Biogeochemistry of Marine Dissolved Organic Matter. Academic Press. ISBN 978-0-12-407153-7.
- ^ Moller, E. F. (2004). "Sloppy feeding in marine copepods: Prey-size-dependent production of dissolved organic carbon". Journal of Plankton Research. 27: 27–35. doi:10.1093/plankt/fbh147.
- ^ Møller, Eva Friis (2007). "Production of dissolved organic carbon by sloppy feeding in the copepods Acartia tonsa, Centropages typicus, and Temora longicornis". Limnology and Oceanography. 52 (1): 79–84. Bibcode:2007LimOc..52...79M. doi:10.4319/lo.2007.52.1.0079.
- ^ Azam, Farooq; Fenchel, Tom; Field, J.G.; Gray, J.S.; Meyer-Reil, L.A.; Thingstad, F. (1983). "The Ecological Role of Water-Column Microbes in the Sea". Marine Ecology Progress Series. 10: 257–263. Bibcode:1983MEPS...10..257A. doi:10.3354/meps010257.
- ^ Van den Meersche, Karel; Middelburg, Jack J.; Soetaert, Karline; van Rijswijk, Pieter; Boschker, Henricus T. S.; Heip, Carlo H. R. (2004). "Carbon-nitrogen coupling and algal-bacterial interactions during an experimental bloom: Modeling a13C tracer experiment". Limnology and Oceanography. 49 (3): 862–878. Bibcode:2004LimOc..49..862V. doi:10.4319/lo.2004.49.3.0862. hdl:1854/LU-434810. ISSN 0024-3590. S2CID 53518458.
- ^ Mentges, A.; Feenders, C.; Deutsch, C.; Blasius, B.; Dittmar, T. (2019). "Long-term stability of marine dissolved organic carbon emerges from a neutral network of compounds and microbes". Scientific Reports. 9 (1): 17780. Bibcode:2019NatSR...917780M. doi:10.1038/s41598-019-54290-z. PMC 6883037. PMID 31780725.
- ^ Wilhelm, Steven W.; Suttle, Curtis A. (1999). "Viruses and nutrient cycles in the sea: viruses play critical roles in the structure and function of aquatic food webs". BioScience. 49 (10): 781–788. doi:10.2307/1313569. JSTOR 1313569.
- ^ Robinson, Carol, and Nagappa Ramaiah. "Microbial heterotrophic metabolic rates constrain the microbial carbon pump." The American Association for the Advancement of Science, 2011.
- ^ Fuhrman, Jed A. (1999). "Marine viruses and their biogeochemical and ecological effects". Nature. 399 (6736): 541–548. Bibcode:1999Natur.399..541F. doi:10.1038/21119. ISSN 0028-0836. PMID 10376593. S2CID 1260399.
- ^ Wigington, Charles H.; Sonderegger, Derek; Brussaard, Corina P. D.; Buchan, Alison; Finke, Jan F.; Fuhrman, Jed A.; Lennon, Jay T.; Middelboe, Mathias; Suttle, Curtis A.; Stock, Charles; Wilson, William H. (March 2016). "Re-examination of the relationship between marine virus and microbial cell abundances". Nature Microbiology. 1 (3): 15024. doi:10.1038/nmicrobiol.2015.24. ISSN 2058-5276. PMID 27572161. S2CID 52829633.
- ^ Tsai, An-Yi, Gwo-Ching Gong, and Yu-Wen Huang. "Importance of the Viral Shunt in Nitrogen Cycling in Synechococcus Spp. Growth in Subtropical Western Pacific Coastal Waters." Terrestrial, Atmospheric & Oceanic Sciences25.6 (2014).
- ^ Anschütz, Anna-Adriana; Maselli, Maira; Traboni, Claudia; Boon, Arjen R.; Stolte, Willem (1 September 2024). "Importance of integrating mixoplankton into marine ecosystem policy and management—Examples from the Marine Strategy Framework Directive". Integrated Environmental Assessment and Management. 20 (5): 1366–1383. doi:10.1002/ieam.4914. hdl:10261/366473. ISSN 1551-3777. Retrieved 5 July 2025. Modified text was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- ^ Lebrato, Mario; Pahlow, Markus; Frost, Jessica R.; Küter, Marie; Jesus Mendes, Pedro; Molinero, Juan-Carlos; Oschlies, Andreas (2019). "Sinking of Gelatinous Zooplankton Biomass Increases Deep Carbon Transfer Efficiency Globally". Global Biogeochemical Cycles. 33 (12): 1764–1783. Bibcode:2019GBioC..33.1764L. doi:10.1029/2019GB006265. Modified text was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- ^ Lebrato, Mario; Pitt, Kylie A.; Sweetman, Andrew K.; Jones, Daniel O. B.; Cartes, Joan E.; Oschlies, Andreas; Condon, Robert H.; Molinero, Juan Carlos & Adler, Laetitia (2012). "Jelly-falls historic and recent observations: a review to drive future research directions". Hydrobiologia. 690 (1): 227–245. doi:10.1007/s10750-012-1046-8. S2CID 15428213.
- ^ Lebrato, M. & Jones, D. O. B. (2009). "Mass deposition event of Pyrosoma atlanticum carcasses off Ivory Coast (West Africa)" (PDF). Limnology and Oceanography. 54 (4): 1197–1209. Bibcode:2009LimOc..54.1197L. doi:10.4319/lo.2009.54.4.1197.
- ^ a b Sweetman, Andrew K. & Chapman, Annelise (2011). "First observations of jelly-falls at the seafloor in a deep-sea fjord". Deep Sea Research Part I: Oceanographic Research Papers. 58 (12): 1206–1211. Bibcode:2011DSRI...58.1206S. doi:10.1016/j.dsr.2011.08.006.
- ^ Burd, Adrian. "Towards a transformative understanding of the ocean's biological pump: Priorities for future research-Report on the NSF Biology of the Biological Pump Workshop" (PDF). OCB: Ocean Carbon & Biogeochemistry. Retrieved 30 October 2016.
- ^ Hays, Graeme C.; Doyle, Thomas K.; Houghton, Jonathan D.R. (2018). "A Paradigm Shift in the Trophic Importance of Jellyfish?". Trends in Ecology & Evolution. 33 (11): 874–884. Bibcode:2018TEcoE..33..874H. doi:10.1016/j.tree.2018.09.001. PMID 30245075. S2CID 52336522.
- ^ a b c d e Roman, Joe; McCarthy, James J. (2010). "The Whale Pump: Marine Mammals Enhance Primary Productivity in a Coastal Basin". PLOS ONE. 5 (10) e13255. Bibcode:2010PLoSO...513255R. doi:10.1371/journal.pone.0013255. PMC 2952594. PMID 20949007. Modified text was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- ^ Brown, Joshua E. (12 October 2010). "Whale poop pumps up ocean health". Science Daily. Retrieved 18 August 2014.
- ^ Longhurst, Alan R.; Glen Harrison, W. (1989). "The biological pump: Profiles of plankton production and consumption in the upper ocean". Progress in Oceanography. 22 (1): 47–123. Bibcode:1989PrOce..22...47L. doi:10.1016/0079-6611(89)90010-4.
- ^ Hutchins, David A.; Wang, Wen-Xiong; Fisher, Nicholas S. (1995). "Copepod grazing and the biogeochemical fate of diatom iron". Limnology and Oceanography. 40 (5): 989–994. Bibcode:1995LimOc..40..989H. doi:10.4319/lo.1995.40.5.0989.
- ^ Steinberg, Deborah K.; Goldthwait, Sarah A.; Hansell, Dennis A. (2002). "Zooplankton vertical migration and the active transport of dissolved organic and inorganic nitrogen in the Sargasso Sea". Deep Sea Research Part I: Oceanographic Research Papers. 49 (8): 1445–1461. Bibcode:2002DSRI...49.1445S. doi:10.1016/S0967-0637(02)00037-7.
- ^ Pershing, Andrew J.; Christensen, Line B.; Record, Nicholas R.; Sherwood, Graham D.; Stetson, Peter B. (2010). "The Impact of Whaling on the Ocean Carbon Cycle: Why Bigger Was Better". PLOS ONE. 5 (8) e12444. Bibcode:2010PLoSO...512444P. doi:10.1371/journal.pone.0012444. PMC 2928761. PMID 20865156.
- ^ "Nature's Solution to Climate Change – IMF F&D". International Monetary Fund. Retrieved 2 November 2019.
- ^ Wiebe, P.H; L.P. Madin; L.R. Haury; G.R. Harbison; L.M. Philbin (1979). "Diel Vertical Migration by Salpa aspera and its potential for large-scale particulate organic matter transport to the deep-sea". Marine Biology. 53 (3): 249–255. Bibcode:1979MarBi..53..249W. doi:10.1007/BF00952433. S2CID 85127670.
- ^ a b c d Jónasdóttir, Sigrún Huld; Visser, André W.; Richardson, Katherine; Heath, Michael R. (29 September 2015). "Seasonal copepod lipid pump promotes carbon sequestration in the deep North Atlantic". Proceedings of the National Academy of Sciences. 112 (39): 12122–12126. doi:10.1073/pnas.1512110112. ISSN 0027-8424. PMC 4593097. PMID 26338976.
- ^ a b c Visser, Andre (2 March 2017). "Calanus hyperboreus and the lipid pump". Limnology and Oceanography. 62 (3): 1155–1165. Bibcode:2017LimOc..62.1155V. doi:10.1002/lno.10492. S2CID 51989153.
- ^ a b c Steinberg, Deborah K.; Landry, Michael R. (3 January 2017). "Zooplankton and the Ocean Carbon Cycle". Annual Review of Marine Science. 9 (1): 413–444. Bibcode:2017ARMS....9..413S. doi:10.1146/annurev-marine-010814-015924. ISSN 1941-1405. PMID 27814033.
- ^ Parent, Genevieve J.; Plourde, Stephane; Turgeon, Julie (1 November 2011). "Overlapping size ranges of Calanus spp. off the Canadian Arctic and Atlantic Coasts: impact on species' abundances". Journal of Plankton Research. 33 (11): 1654–1665. doi:10.1093/plankt/fbr072. ISSN 0142-7873.
- ^ Kristiansen, Inga; Gaard, Eilif; Hátún, Hjálmar; Jónasdóttir, Sigrún; Ferreira, A. Sofia A. (1 May 2016). "Persistent shift of Calanus spp. in the southwestern Norwegian Sea since 2003, linked to ocean climate". ICES Journal of Marine Science. 73 (5): 1319–1329. doi:10.1093/icesjms/fsv222. ISSN 1054-3139.
- ^ Jensen, Maj Holst; Nielsen, Torkel Gissel; Dahllöf, Ingela (28 April 2008). "Effects of pyrene on grazing and reproduction of Calanus finmarchicus and Calanus glacialis from Disko Bay, West Greenland". Aquatic Toxicology. 87 (2): 99–107. Bibcode:2008AqTox..87...99J. doi:10.1016/j.aquatox.2008.01.005. ISSN 0166-445X. PMID 18291539.
- ^ a b c d Tanet, Lisa; Martini, Séverine; Casalot, Laurie; Tamburini, Christian (2020). "Reviews and syntheses: Bacterial bioluminescence – ecology and impact in the biological carbon pump". Biogeosciences. 17 (14): 3757–3778. Bibcode:2020BGeo...17.3757T. doi:10.5194/bg-17-3757-2020. Modified text was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- ^ Kayler, Z., Janowiak, M., Swanston, C. (2017). "The Global Carbon Cycle". Considering Forest and Grassland Carbon in Land Management. Vol. 95. United States Department of Agriculture, Forest Service. pp. 3–9. doi:10.2737/WO-GTR-95.
{{cite book}}:|journal=ignored (help)CS1 maint: multiple names: authors list (link) - ^ NASA (16 June 2011). "The Slow Carbon Cycle". Archived from the original on 16 June 2012. Retrieved 24 June 2012.
- ^ Marinov, I.; Gnanadesikan, A.; Toggweiler, J. R.; Sarmiento, J. L. (2006). "The Southern Ocean biogeochemical divide". Nature. 441 (7096): 964–967. Bibcode:2006Natur.441..964M. doi:10.1038/nature04883. PMID 16791191. S2CID 4428683.
- ^ Takahashi, Taro; Sutherland, Stewart C.; Sweeney, Colm; Poisson, Alain; Metzl, Nicolas; Tilbrook, Bronte; Bates, Nicolas; Wanninkhof, Rik; Feely, Richard A.; Sabine, Christopher; Olafsson, Jon; Nojiri, Yukihiro (2002). "Global sea–air CO2 flux based on climatological surface ocean pCO2, and seasonal biological and temperature effects". Deep Sea Research Part II: Topical Studies in Oceanography. 49 (9–10): 1601–1622. Bibcode:2002DSRII..49.1601T. doi:10.1016/S0967-0645(02)00003-6.
- ^ Orr, J. C., E. Maier-Reimer, U. Mikolajewicz, P. Monfray, J. L. Sarmiento, J. R. Toggweiler, N. K. Taylor, J. Palmer, N. Gruber, C. L. Sabine, C. Le Quéré, R. M. Key and J. Boutin (2001). Estimates of anthropogenic carbon uptake from four three-dimensional global ocean models. Global Biogeochem. Cycles 15, 43–60.
- ^ "Study reveals uncertainty in how much carbon the ocean absorbs over time". MIT News | Massachusetts Institute of Technology. 5 April 2021. Retrieved 7 May 2021.
- ^ Riebesell, U., Schulz, K.G., Bellerby, R.G.J., Botros, M., Fritsche, P., Meyerhöfer, M., Neill, C., Nondal, G., Oschlies, A., Wohlers, J. and Zöllner, E. (2007). Enhanced biological carbon consumption in a high CO2 ocean. Nature 450, 545–548.
- ^ a b Parmentier, Frans-Jan W.; Christensen, Torben R.; Rysgaard, Søren; Bendtsen, Jørgen; Glud, Ronnie N.; Else, Brent; Van Huissteden, Jacobus; Sachs, Torsten; Vonk, Jorien E.; Sejr, Mikael K. (2017). "A synthesis of the arctic terrestrial and marine carbon cycles under pressure from a dwindling cryosphere". Ambio. 46 (Suppl 1): 53–69. Bibcode:2017Ambio..46S..53P. doi:10.1007/s13280-016-0872-8. PMC 5258664. PMID 28116680. Modified text was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- ^ a b Parmentier, Frans-Jan W.; Christensen, Torben R.; Sørensen, Lise Lotte; Rysgaard, Søren; McGuire, A. David; Miller, Paul A.; Walker, Donald A. (2013). "The impact of lower sea-ice extent on Arctic greenhouse-gas exchange". Nature Climate Change. 3 (3): 195–202. Bibcode:2013NatCC...3..195P. doi:10.1038/nclimate1784.
- ^ Orr, James C.; Fabry, Victoria J.; Aumont, Olivier; Bopp, Laurent; Doney, Scott C.; Feely, Richard A.; Gnanadesikan, Anand; Gruber, Nicolas; Ishida, Akio; et al. (2005). "Anthropogenic ocean acidification over the twenty-first century and its impact on calcifying organisms" (PDF). Nature. 437 (7059): 681–686. Bibcode:2005Natur.437..681O. doi:10.1038/nature04095. PMID 16193043. S2CID 4306199.
- ^ Armstrong, Robert A.; Lee, Cindy; Hedges, John I.; Honjo, Susumu & Wakeham, Stuart G. (2001). "A new, mechanistic model for organic carbon fluxes in the ocean based on the quantitative association of POC with ballast minerals". Deep Sea Research Part II: Topical Studies in Oceanography. 49 (1–3): 219–236. Bibcode:2001DSRII..49..219A. doi:10.1016/s0967-0645(01)00101-1.
- ^ Bopp, L.; Aumont, O.; Cadule, P.; Alvain, S.; Gehlen, M. (2005). "Response of diatoms distribution to global warming and potential implications: A global model study". Geophysical Research Letters. 32 (19): n/a. Bibcode:2005GeoRL..3219606B. doi:10.1029/2005GL023653.
- ^ Iglesias-Rodriguez, M. D.; Halloran, P. R.; Rickaby, R. E. M.; Hall, I. R.; Colmenero-Hidalgo, E.; Gittins, J. R.; Green, D. R. H.; Tyrrell, T.; Gibbs, S. J.; von Dassow, P.; Rehm, E.; Armbrust, E. V.; Boessenkool, K. P. (2008). "Phytoplankton Calcification in a High-CO2 World". Science. 320 (5874): 336–340. Bibcode:2008Sci...320..336I. doi:10.1126/science.1154122. PMID 18420926. S2CID 206511068.
- ^ Rivero-Calle, Sara; Gnanadesikan, Anand; Castillo, Carlos E. Del; Balch, William M. & Guikema, Seth D. (2015). "Multidecadal increase in North Atlantic coccolithophores and the potential role of rising CO2". Science. 350 (6267): 1533–1537. Bibcode:2015Sci...350.1533R. doi:10.1126/science.aaa8026. PMID 26612836.
- ^ Jensen, Lasse Ørsted; Mousing, Erik Askov; Richardson, Katherine (2017). "Using species distribution modelling to predict future distributions of phytoplankton: Case study using species important for the biological pump". Marine Ecology. 38 (3) e12427. Bibcode:2017MarEc..38E2427J. doi:10.1111/maec.12427.
- ^ Petrou, Katherina; Nielsen, Daniel (27 August 2019). "Acid oceans are shrinking plankton, fueling faster climate change". phys.org. Retrieved 7 September 2019.
- ^ Siegel, David A.; Buesseler, Ken O.; Behrenfeld, Michael J.; Benitez-Nelson, Claudia R.; Boss, Emmanuel; Brzezinski, Mark A.; Burd, Adrian; Carlson, Craig A.; D'Asaro, Eric A.; Doney, Scott C.; Perry, Mary J.; Stanley, Rachel H. R.; Steinberg, Deborah K. (8 March 2016). "Prediction of the Export and Fate of Global Ocean Net Primary Production: The EXPORTS Science Plan". Frontiers in Marine Science. 3. Frontiers Media SA. doi:10.3389/fmars.2016.00022. hdl:1912/8770. ISSN 2296-7745.
- ^ Committee on Earth Observation Satellites (2014) "CEOS strategy for carbon observations from space". Response to the Group on Earth Observations (GEO) carbon strategy. Printed in Japan by JAXA and I&A Corporation.
- ^ a b c d e Levin, L.A., Bett, B.J., Gates, A.R., Heimbach, P., Howe, B.M., Janssen, F., McCurdy, A., Ruhl, H.A., Snelgrove, P., Stocks, K.I., Bailey, D. and 27 others (2019) "Global observing needs in the deep ocean". Frontiers in Marine Science, 6: 241. doi:10.3389/fmars.2019.00241. Modified text was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
| Groups |  | |
|---|---|---|
| Microbiology | ||
| Motion | ||
| Ecology | ||
| Plants | ||
| Marine | ||
| Human related | ||
| Techniques | ||
| Other | ||
Biological pump
View on GrokipediaOverview
Definition and Core Mechanism
The biological carbon pump, also known as the biological pump, encompasses the biological, physical, and chemical processes that actively transport organic carbon from the ocean's sunlit surface layer to deeper waters, thereby sequestering it from rapid exchange with the atmosphere. This mechanism lowers the partial pressure of CO2 in surface seawater relative to the atmosphere, facilitating oceanic uptake of atmospheric carbon dioxide. Primary producers, predominantly phytoplankton, initiate the process by fixing dissolved inorganic carbon—primarily bicarbonate and CO2—into organic biomass through photosynthesis, utilizing sunlight and macronutrients like nitrate and phosphate.[8][2] At its core, the pump operates via the export of a fraction of this fixed carbon below the euphotic zone, typically defined as the depth of the annual maximum mixed layer or around 100-200 meters. The export ratio, or e-ratio, represents the proportion of net primary production that escapes surface remineralization and sinks as particulate organic carbon (POC), often in forms such as zooplankton fecal pellets, phytoplankton aggregates, or dead cells. These sinking particles, collectively termed marine snow, undergo gradual remineralization by heterotrophic bacteria and protists in the mesopelagic zone, releasing dissolved inorganic carbon that accumulates in deep waters due to sluggish vertical mixing and the ocean's thermal stratification. This vertical partitioning sustains a gradient in dissolved inorganic carbon concentrations, with surface values around 2,000 μmol kg⁻¹ and deep values exceeding 2,200 μmol kg⁻¹ in many basins.[9][10] The efficiency of carbon sequestration depends on the balance between export flux and remineralization depth; particles that sink beyond the permanent thermocline contribute to long-term storage on millennial timescales, while shallower remineralization returns carbon more quickly to the surface. Globally, the biological pump is estimated to export 5-12 GtC per year from the surface ocean, representing 10-25% of net primary production, though regional variations are pronounced due to nutrient availability, food web structure, and particle ballasting by minerals like calcium carbonate or opal. Dissolved organic carbon also plays a role, with a fraction exported via physical mixing or microbial degradation gradients, but the particulate pathway dominates vertical flux.[9][2]Role in the Global Carbon Cycle
The biological pump facilitates the export of organic carbon from the sunlit surface ocean to the interior, where remineralization occurs slowly, thereby sustaining a vertical gradient in dissolved inorganic carbon (DIC) and regulating atmospheric CO₂ levels on timescales from decades to millennia.[11] This process complements the solubility pump by enabling deeper and longer-term carbon storage through biological mediation rather than solely physical dissolution.[11] Globally, phytoplankton primary production fixes approximately 50 Pg C yr⁻¹, with 1–40% potentially available for export depending on regional productivity and food web dynamics.[12] Export fluxes at the base of the euphotic zone or ~100 m depth are estimated at 5–12 Pg C yr⁻¹ for particulate organic carbon, though top-down modeling from multidecadal hydrographic data yields a total organic carbon flux of 15.0 ± 1.1 Pg C yr⁻¹, including 10.6 ± 0.1 Pg C yr⁻¹ via sinking particles and vertical migration.[11][13] Transfer efficiency decreases with depth due to remineralization, such that only ~10–20% of exported carbon reaches 1,000 m, resulting in net sequestration of 0.2–1.0 Pg C yr⁻¹ into the deep ocean and sediments on centennial scales.[11][12] This sequestration contributes to an atmospheric CO₂ drawdown of ~0.5–1.0 Pg C yr⁻¹ by enhancing the ocean's DIC inventory and isolation from the atmosphere, far exceeding the solubility pump's role in long-term storage (~2 Pg C yr⁻¹ annual uptake but shallower).[11] Regional variations, such as higher export in nutrient-rich upwelling zones, amplify the pump's global impact, while microbial degradation in the mesopelagic zone modulates flux attenuation, with power-law decay exponents (b) ranging from 0.2 in oligotrophic gyres to 1.0 in eutrophic regions.[13][12] Uncertainties in these estimates arise from sparse observations and model assumptions about remineralization rates, underscoring the need for continued empirical validation.[11]Carbon Forms and Oceanic Pools
Organic Carbon: Dissolved and Particulate
In the ocean, organic carbon exists primarily in two forms: particulate organic carbon (POC), which consists of particles larger than 0.7 micrometers, and dissolved organic carbon (DOC), which includes molecules passing through a 0.7-micrometer filter.[9] These forms arise from phytoplankton primary production, where net primary production (NPP) of approximately 50 gigatons of carbon per year is partitioned, with roughly 10-20% entering the particulate pool suitable for sinking and the remainder initially dissolved or rapidly converted.[14] POC drives the core vertical flux of the biological pump by aggregating into sinking particles like marine snow, enabling export from the euphotic zone to depths below 100 meters at rates of 5-10 gigatons of carbon annually.[15] In contrast, DOC, comprising a vast reservoir of 600-700 gigatons of carbon—over 200 times the biomass—predominantly remains in the surface ocean or is remineralized by microbes, contributing less directly to deep sequestration unless transformed into refractory forms.[16] POC originates mainly from phytoplankton exudates, zooplankton fecal pellets, and microbial detritus, with export efficiency varying by region and ecosystem structure; in high-nutrient low-chlorophyll areas, POC fluxes can exceed 1 gram of carbon per square meter per day at 100 meters depth.[17] Sinking POC remineralizes with depth following a power-law decay, where 1-2% of surface NPP reaches the seafloor, sequestering carbon for centuries to millennia in sediments.[18] Ballasted particles, such as those coated with biogenic minerals, enhance sinking speeds to 10-100 meters per day, amplifying the pump's efficiency compared to unballasted organic aggregates.[19] Observational data from sediment traps and thorium-234 proxies confirm POC as the dominant vector for biological pump-mediated carbon drawdown, though its flux attenuates rapidly due to solubilization and grazing.[8] DOC, largely produced via phytoplankton leakage (2-50% of fixed carbon) and sloppy feeding, partitions into labile (rapidly consumed) and refractory (long-lived) pools, with the latter persisting for millennia through microbial processing known as the microbial carbon pump.[20] [12] While DOC export via subduction or mixing contributes about 0.48 petagrams of carbon per year to the deep ocean—comparable to POC in some models—its role in sequestration is indirect, as physical injection rather than biological sinking dominates, and much returns to dissolved inorganic carbon via respiration.[21] In regions like the Southern Ocean, DOC fluxes may rival POC for total organic carbon export, but global sequestration favors POC due to gravitational settling over diffusive DOC transport.[22] Refractory DOC accumulation, however, buffers atmospheric CO2 by resisting degradation, with bacterial communities transforming 20-50% of labile inputs into persistent forms annually.[23] The interplay between POC and DOC influences pump efficiency; high DOC release from blooms reduces direct POC export but sustains microbial loops that regenerate nutrients and indirectly support further production.[24] Empirical assessments indicate that while POC handles the "active" sinking component, DOC's "passive" persistence—estimated at 0.2-0.5 gigatons sequestered yearly—extends the pump's temporal scale, though vulnerabilities to warming-induced remineralization could diminish both pools' contributions. Quantifying these fluxes remains challenging, with satellite-derived NPP and in situ profiles revealing regional variability, such as elevated POC export in iron-fertilized waters versus DOC dominance in stratified gyres.[8]Inorganic Carbon and Mineral Forms
Particulate inorganic carbon (PIC) in the biological pump refers to biogenic calcium carbonate (CaCO3) particles exported from the surface ocean to deeper layers through sinking.[8] These particles form via calcification by planktonic organisms, which precipitate CaCO3 from seawater's dissolved inorganic carbon pool, primarily bicarbonate ions.[3] Calcification releases CO2 locally, but when coupled with photosynthetic carbon fixation in the same organisms, the net effect on surface ocean CO2 can vary; export of PIC contributes to long-term sequestration by isolating it from surface waters until potential dissolution at depth.[25] The primary mineral forms of oceanic PIC are calcite and aragonite, two polymorphs of CaCO3 differing in crystal structure, solubility, and biological producers.[26] Calcite, the more stable and less soluble form, dominates PIC export and is produced by coccolithophores (e.g., Emiliania huxleyi and Coccolithus pelagicus) as intricate scales called coccoliths, and by planktonic foraminifera as chambered tests.[8] Aragonite, more soluble and prone to dissolution in undersaturated waters, is secreted by pteropods and some benthic foraminifera as thin shells.[27] Global PIC production is estimated at 0.8–1.4 gigatons of carbon per year, with coccolithophores contributing the majority in open ocean gyres.[28] PIC particles enhance the efficiency of the biological pump by acting as ballast, increasing sinking rates of associated organic matter through density and aggregation effects.[8] However, PIC solubility increases with depth due to decreasing carbonate saturation states, leading to dissolution that regenerates dissolved inorganic carbon (DIC) in the deep ocean without returning it to the atmosphere on millennial timescales.[26] Aragonite dissolves at shallower depths (typically above 500–1000 meters in many regions) compared to calcite (below 3000–4000 meters), influencing the vertical distribution of carbon remineralization.[29] Ocean acidification, driven by anthropogenic CO2 uptake, reduces carbonate ion availability, impairing calcification and PIC export while accelerating dissolution, potentially weakening this component of the pump.[30]Primary Production and Export
Phytoplankton Dynamics
Phytoplankton, consisting mainly of eukaryotic algae and prokaryotic cyanobacteria, drive oceanic primary production by photosynthetically fixing dissolved inorganic carbon into particulate and dissolved organic forms, contributing 39–58 Gt C yr⁻¹ of net primary production globally.[31] This process, occurring predominantly in the euphotic zone (upper ~100–200 m), establishes the initial organic carbon pool for the biological pump, with fixation rates varying by latitude, season, and nutrient regime. Picophytoplankton (<2 μm) account for ~39% of total production (~19 Gt C yr⁻¹), dominating oligotrophic subtropical gyres, while larger nano- (2–20 μm) and microplankton (>20 μm) prevail in nutrient-enriched upwelling zones and high-latitude blooms.[32] Population dynamics hinge on resource availability and physical forcing: macronutrients (nitrogen as nitrate/ammonium, phosphorus as phosphate, silicate for diatoms) limit growth in ~70% of ocean surface waters, while iron constrains high-nutrient low-chlorophyll (HNLC) regions like the Southern Ocean and equatorial Pacific, reducing fixation by up to 50% without supplementation.[33] Light intensity and photoperiod regulate division rates (typically 0.1–2 doublings day⁻¹), with photoinhibition above ~500 μmol photons m⁻² s⁻¹, and temperature optima shifting from ~10°C in polar species to 25–30°C in tropical ones, influencing metabolic efficiency and carbon allocation to biomass versus exudates. Water column stratification suppresses vertical nutrient flux, favoring small cells with high surface-to-volume ratios, whereas mixing events or upwelling elevate productivity by 5–10-fold, often culminating in blooms where biomass exceeds 10 mg chl-a m⁻³.[34] Bloom formation, a hallmark of phytoplankton dynamics, pulses carbon export by aggregating cells into fast-sinking aggregates or marine snow, with diatom-dominated events in temperate/polar springs exporting 5–10 Gt C yr⁻¹ globally via the pump.[35] Succession patterns—initial nutrient exploitation by fast-growing diatoms yielding to nutrient-specialists like dinoflagellates or prymnesiophytes—alter export potential, as silica-ballasted diatoms sink at 10–100 m day⁻¹ versus <1 m day⁻¹ for unballasted flagellates. Microplankton blooms yield higher export ratios (export/NPP = 10–25%) than pico/nano-dominated regimes (<5%), due to larger particle formation and reduced remineralization in surface layers.[36] Certain taxa exhibit active vertical migration (10–100 m daily), accessing deep nutrients to sustain fixation in stratified oligotrophic waters, contributing up to 40% of new production (~28 Tg N yr⁻¹ equivalent) and directly enhancing export by 25% through fecal pellet production or direct sinking.[37] Community shifts under warming or acidification—e.g., toward smaller cells or non-silicifiers—could diminish pump efficiency by 10–20%, as evidenced in mesocosm experiments and models linking size structure to flux attenuation.[36] Empirical data from sediment traps and thorium-234 proxies confirm that bloom termination, via grazing or nutrient depletion, triggers 50–80% of annual export in productive ecosystems, underscoring the pulsed nature of phytoplankton-driven carbon transfer.[38]Initial Carbon Fixation and Export Flux
Initial carbon fixation in the biological pump refers to the photosynthetic conversion of dissolved inorganic carbon (DIC), primarily bicarbonate and CO2, into organic carbon by marine phytoplankton in the euphotic zone. This process yields net primary production (NPP) estimated at 49-50 gigatons of carbon (GtC) annually across global ocean ecosystems.[39] [40] Phytoplankton groups such as diatoms, dinoflagellates, coccolithophores, and cyanobacteria perform this fixation, with efficiency governed by light intensity, nutrient concentrations (e.g., nitrate, phosphate, iron), and temperature.[17] The reaction fundamentally follows 106 CO2 + 16 HNO3 + H3PO4 + 122 H2O → (CH2O)106(NH3)16H3PO4 + 138 O2, representing the Redfield ratio stoichiometry observed in marine organic matter.[10] Export flux quantifies the portion of fixed organic carbon that sinks out of the surface mixed layer or euphotic zone as particulate organic carbon (POC), evading rapid remineralization by herbivores and microbes. Globally, this flux at approximately 75 meters depth is estimated at 15.00 ± 1.12 GtC per year, corresponding to an export ratio (e-ratio) of POC export divided by NPP of roughly 0.3.[13] The e-ratio varies spatially and temporally, typically ranging from 0.05 in oligotrophic gyres to over 0.4 in high-nutrient, low-chlorophyll (HNLC) regions, influenced by factors like grazing pressure, particle aggregation, and seasonal stratification.[41] [9] Export initiates via mechanisms such as phytoplankton senescence, fecal pellet production from zooplankton grazing, and the formation of marine snow aggregates, which enhance sinking rates beyond those of individual cells.[17] Measurement of initial fixation relies on satellite-derived chlorophyll estimates calibrated against in situ NPP assays, while export flux is assessed through sediment traps, thorium-234 proxies, and neutrally buoyant floats, revealing discrepancies due to methodological assumptions and regional biases.[42] Recent inverse modeling from multidecadal hydrographic data confirms the 15 GtC export magnitude, underscoring the pump's role in sequestering about 30% of fixed carbon below the euphotic zone before substantial respiration.[13] Variations in export efficiency link to ecosystem structure, with diatom-dominated blooms fostering higher fluxes than picophytoplankton assemblages due to faster sinking of siliceous tests.[9]Particle Formation and Sinking Processes
Marine Snow and Aggregation
Marine snow consists of large aggregates (>500 µm) of organic detritus, including decaying phytoplankton, fecal material, and microbial biofilms, that form in the ocean's surface waters and sink to deeper layers, facilitating carbon export in the biological pump.[43] These particles, often termed "ocean snowflakes," represent a primary mechanism for transferring particulate organic carbon (POC) from the euphotic zone to the mesopelagic and bathypelagic depths.[12] Aggregation processes drive their formation, where smaller colloids, bacteria, and phytoplankton cells collide and adhere via biophysical and biochemical mechanisms, such as sticky exopolymers like transparent exopolymer particles (TEP).[44] Particle aggregation is enhanced by turbulence in the upper ocean, with studies indicating optimal formation at turbulent kinetic energy dissipation rates below 10^{-6} W kg^{-1}, beyond which shear forces disaggregate the flocs.[45] Biopolymers secreted by phytoplankton and bacteria act as glues, promoting coagulation; for instance, TEP concentration correlates positively with aggregation efficiency, increasing sinking velocities from millimeters to centimeters per day.[46] In situ observations in regions like the Black Sea and Gulf of Mexico reveal marine snow settling speeds ranging from 10 to 300 m day^{-1}, depending on aggregate density and size, with larger, stringy morphotypes exhibiting higher export potential than compact forms.[47][43] Recent research highlights interactions with biogels, such as polysaccharide gels, that adhere to sinking particles, reducing their velocities by up to 45% within a day through increased drag and buoyancy, thereby modulating remineralization depth and carbon sequestration efficiency.[48] Marine snow contributes significantly to POC flux, often comprising 20-50% of exported carbon in productive systems, though fragmentation from zooplankton grazing or physical disaggregation can recycle up to 70% of aggregates back to dissolved forms before deep burial.[49][50] These dynamics underscore marine snow's role as a variable but critical conduit in the biological pump, influenced by hydrodynamic and biological forcings.[18]Ballast Effects and Biomineralization
Biomineralization in marine ecosystems involves the production of dense inorganic structures, primarily calcium carbonate (CaCO3) by coccolithophores and foraminifera, and biogenic silica (BSi or opal) by diatoms, which contribute to the biological carbon pump by facilitating the export of particulate organic carbon (POC).[51] These minerals form protective shells or frustules, with densities significantly higher than surrounding seawater (CaCO3 at approximately 2.7 g/cm³ and opal at 2.1–2.3 g/cm³ compared to seawater's 1.025 g/cm³).[52] The ballast effect refers to the enhanced sinking rates of organic particles due to association with these biogenic minerals, which increase particle density and reduce remineralization in the upper ocean, thereby promoting deeper carbon sequestration.[53] According to the ballast hypothesis proposed by Armstrong et al. (2002), a substantial portion of deep-sea POC flux correlates with mineral content, with calcite ballasting approximately one-third and opal one-sixth of exported POC in many ocean regions.[54] Empirical data from sediment traps indicate that sinking velocities of marine snow aggregates can increase by factors of 2–10 with higher ballast ratios, as minerals counteract the buoyancy of low-density organic matter.[55] In the Southern Ocean, biogenic minerals dominate sinking particle mass, comprising over 85% as opal and CaCO3 at depths around 1000 m, underscoring their role in efficient export despite high remineralization rates elsewhere.[52] However, spatial variability exists; for instance, in lithogenic-poor regions, the ballast effect is primarily biogenic, while riverine inputs enhance it in marginal seas like the Bay of Bengal.[56] Biomineralization also links to carbon fixation, as calcification by coccolithophores co-occurs with photosynthesis, though net alkalinity effects can modulate CO2 drawdown.[57] Despite these benefits, ballast efficiency is modulated by particle size, aggregation, and environmental factors; smaller particles with low mineral content sink slowly, limiting export, while dissolution of CaCO3 in undersaturated deep waters can release associated POC to remineralization. Overall, biogenic minerals are estimated to enhance global POC export by 20–50% through ballasting, based on flux attenuation models.[58]Fecal Pellets and Sloppy Feeding
Zooplankton, particularly mesozooplankton such as copepods and krill, produce fecal pellets during grazing on phytoplankton and detrital particles, repackaging small, slow-sinking organic matter into dense, cylindrical or oval structures that sink rapidly at velocities often exceeding 100–500 meters per day.[59] [60] These pellets facilitate efficient carbon export in the biological pump by minimizing remineralization in the upper ocean, with contributions to total particulate organic carbon (POC) flux reaching up to 100% in high-latitude regions like the Arctic and Antarctic, and 20–50% in subtropical gyres.[9] [61] Pellet carbon content varies by producer; for instance, krill and salp pellets in the Southern Ocean can comprise 40–70% organic carbon by dry weight, enhancing sequestration when they reach depths below 1,000 meters before significant degradation.[60] [62] Fecal pellet sinking efficiency depends on factors including pellet size (typically 50–500 micrometers in length), mineral ballasting from ingested coccoliths or siliceous debris, and environmental conditions like temperature, which influence remineralization rates.[63] [64] In the South China Sea, zooplankton fecal pellets have been observed to dominate passive POC flux at depths of 200–1,000 meters, with sinking speeds supporting transfer to the deep ocean and contributing to long-term carbon storage.[65] However, pellets often attenuate rapidly with depth due to fragmentation or microbial degradation, with long, intact forms comprising less than 5% of flux below 1,000 meters in some open-ocean settings, underscoring the need for integrated flux measurements combining traps and optical sensors.[66] [67] Sloppy feeding occurs when zooplankton handling of prey causes physical breakage or leakage, releasing 10–40% of grazed carbon as dissolved organic matter (DOM) or small particulate fragments rather than fully ingested material.[68] [69] These by-products contribute indirectly to the biological pump through aggregation into marine snow or uptake by bacteria, which may form sinking aggregates, though much of the DOM remains in the surface mixed layer, promoting nutrient recycling over export.[70] [71] In contrast to compact fecal pellets, sloppy feeding fragments sink more slowly (often <10 meters per day) and are prone to disaggregation, reducing their net contribution to deep carbon flux; models estimate sloppy feeding accounts for 20% of grazing-derived small POC in some simulations, but empirical data highlight its role in surface-layer retention rather than sequestration.[72] [73] The interplay between fecal pellets and sloppy feeding modulates pump efficiency, as higher grazing pressures increase both pellet production and DOM release, with community composition—e.g., dominance by gelatinous zooplankton like salps favoring larger, faster-sinking pellets—altering export ratios.[74] [75] Recent meta-analyses confirm fecal pellets as a primary vector for POC transfer, while sloppy feeding primarily supports the microbial loop, emphasizing zooplankton mediation in partitioning carbon between recycling and export pathways.[76][77]Heterotrophic and Remineralization Processes
Zooplankton Grazing and Vertical Migrations
Zooplankton grazing on phytoplankton repackages photosynthetically fixed carbon into fecal pellets, which sink more rapidly than uneaten phytoplankton cells, thereby enhancing the efficiency of carbon export to the deep ocean. These pellets, often dense and aggregated, can achieve sinking speeds of hundreds of meters per day, facilitating the transfer of particulate organic carbon (POC) below the euphotic zone and contributing variably to total export fluxes, such as 3.4% at 500 m depth in the northern South China Sea. Grazing also occurs through "sloppy feeding," where fragmented prey release dissolved organic carbon (DOC) and small particles, some of which aggregate into marine snow, though this process may reduce net export if remineralization dominates. In regions like the Kerguelen Plateau, mesozooplankton grazing significantly influences export regimes by controlling phytoplankton biomass and directing carbon toward sinking pathways. However, grazing rates introduce substantial uncertainty in models of the biological pump, as they mediate both trophic transfer to higher levels and deep sequestration.[62] Diel vertical migration (DVM) by zooplankton, involving nocturnal ascent to surface waters for feeding and diurnal descent to deeper layers for predator avoidance, actively transports carbon downward through migrating biomass, fecal pellet release, and respiration at depth. This process can account for up to 27% of POC export below the mixed layer in the North Atlantic, with modeled increases in surface export flux ranging from 16-30% attributable to DVM. Migrants, including copepods and euphausiids, carry fixed carbon in their bodies and excrete labile material in mesopelagic zones, where remineralization is slower, effectively amplifying the biological pump's sequestration potential. In the Canary Current, zooplankton DVM contributes measurable respiratory carbon flux, underscoring its role in regional export dynamics. Recent assessments confirm DVM as a globally ubiquitous mechanism, with deep-sea biomass surveys indicating substantial vertical carbon flux via this "ladder" of migrations across trophic levels.[78][79][80]Microbial Loop and Bacterial Roles
The microbial loop describes the rapid cycling of dissolved organic matter (DOM) in marine surface waters, where phytoplankton-derived DOM is primarily taken up by heterotrophic bacteria, fueling bacterial growth and subsequent grazing by protists. This process, first conceptualized in the 1980s, recycles carbon and nutrients within the euphotic zone, with bacteria assimilating labile DOM fractions—such as amino acids, sugars, and low-molecular-weight compounds—at rates that can exceed 50% of primary production in oligotrophic systems.[81] Bacterial growth efficiency typically ranges from 10-40%, meaning a substantial portion of assimilated carbon is respired as CO₂, limiting transfer to higher trophic levels or export.[12] In the biological pump, the microbial loop acts as a retention mechanism, remineralizing organic carbon to dissolved inorganic carbon (DIC) before it can aggregate into sinking particles, thereby reducing export flux to the deep ocean. Studies indicate that microbial remineralization consumes 20-50% of sinking particulate organic carbon (POC) in the upper 100-200 meters, with efficiency varying by temperature, nutrient availability, and DOM quality; warmer conditions enhance bacterial respiration rates, potentially decreasing pump efficiency by up to 15% per degree Celsius increase.[82] [18] This remineralization is depth-dependent, with higher rates in the mesopelagic zone where attached bacteria degrade ~10-30% of POC flux, influenced by particle size and ballast minerals.[83] Bacteria also contribute to carbon export indirectly through attachment to marine snow or fecal pellets, forming biofilms that can either accelerate remineralization or enhance sinking via increased density. In DOM-dominated pathways, bacterial processing generates refractory DOM, which evades rapid degradation and supports a complementary microbial carbon pump, sequestering an estimated 0.2-1 Gt C year⁻¹ in the deep ocean as persistent compounds.[84] However, the net effect of bacterial roles often favors surface retention, as evidenced by global models showing microbial loops diverting up to 70% of fixed carbon from export in low-productivity gyres.[85] Empirical data from sediment traps and incubation experiments confirm these dynamics, highlighting bacteria's dual role in both cycling and selective preservation of ocean carbon.[86]Viral Shunt and Lysis
The viral shunt describes the ecological process in marine ecosystems where viruses infect and lyse prokaryotic and eukaryotic microbes, releasing intracellular organic contents as dissolved organic matter (DOM) that bacteria rapidly assimilate and remineralize, thereby recycling carbon within the upper ocean rather than permitting its export as sinking particulate organic carbon (POC).[87] This pathway, distinct from zooplankton grazing which can produce exportable fecal pellets, attenuates the biological carbon pump's efficiency by shunting fixed carbon back into microbial loops and promoting respiration over sequestration.[88] Viral lysis thus sustains high rates of nutrient turnover in the euphotic zone, potentially enhancing primary production through recycled nutrients but limiting deep-ocean carbon burial.[87] Marine viruses dominate numerically, with abundances often 10-fold higher than bacterial hosts—reaching up to 22 × 10⁹ particles per liter in tropical surface waters—and daily infection rates impacting 20–40% of microbes in the euphotic zone.[89] [87] Infection culminates in host cell lysis after viral replication, with typical burst sizes (viral particles released per lysed cell) averaging around 45, though varying by virus-host pair and environmental conditions.[90] The resultant DOM pulse—comprising labile compounds like amino acids and nucleotides—fuels bacterial secondary production, evidenced by correlations between viral abundance and bacterial specific growth rates with 3–6 hour lags in field observations.[87] Quantitatively, viral lysis imposes substantial mortality on microbial biomass; in the Southern Ocean, it accounted for 58% of seasonal phytoplankton carbon losses (approximately 2674 µg C L⁻¹ out of 4645 µg C L⁻¹ total), with lysis rates (0.29 d⁻¹) comparable to protistan grazing (0.31 d⁻¹).[88] In tropical oligotrophic systems, virus-mediated bacterial mortality rates span 0.11–1.71 mg C m⁻³ hour⁻¹ (mean 0.45 mg C m⁻³ hour⁻¹), contributing to estimated global releases of ~145 Gt C yr⁻¹ from lysis in such waters.[87] These fluxes redirect carbon from higher trophic transfer, as lysed cells bypass grazer-mediated sinking, and may alter export efficiency based on eukaryotic virus dominance, where lytic communities correlate with reduced carbon export to depth.31199-8) [88]Additional Biological Pathways
Macroorganisms and Whale Pump
Macroorganisms, including large marine vertebrates such as whales, seals, and fish, contribute to the biological carbon pump by facilitating nutrient recycling, producing sinking biogenic particles, and enabling vertical carbon transport through their behaviors and life cycles. These organisms often graze on prey in nutrient-rich zones and excrete waste or die in surface waters, redistributing limiting nutrients like iron and nitrogen to stimulate phytoplankton growth, which in turn enhances primary production and subsequent export of organic carbon to depth. Unlike smaller planktonic contributors, macroorganisms influence the pump on larger spatial scales due to their migrations and high biomass turnover, though their overall impact remains subordinate to microbial and zooplankton processes in global flux estimates.[91] The whale pump specifically describes the mechanism by which cetaceans, particularly baleen whales, amplify carbon sequestration. Whales consume iron-replete krill and small fish in polar or deep foraging grounds, then migrate to oligotrophic surface waters where they release iron-enriched fecal plumes, alleviating iron limitation in high-nutrient, low-chlorophyll (HNLC) regions and promoting phytoplankton blooms that fix atmospheric CO2. This nutrient shuttling mirrors aspects of the microbial loop but operates via macro-scale animal migration, with studies in the Gulf of Maine estimating that whale-mediated iron recycling could support up to 20% of local primary productivity through recycled nitrogen and carbon. Fecal matter from whales also aggregates into marine snow, accelerating particulate organic carbon (POC) sinking rates compared to smaller zooplankton pellets.[92][93] Additional contributions from whale carcasses, known as whale falls, provide direct sequestration as massive carbon-rich bodies sink to the seafloor, supporting chemosynthetic communities and burying organic matter beyond remineralization zones; a blue whale carcass can sequester approximately 30-50 tons of carbon for decades. Historical industrial whaling depleted whale populations by over 90% in some basins, reducing this pump's efficiency, with models suggesting full recovery of Southern Ocean blue and fin whale stocks could enhance sequestration by 1.6 million tons of CO2 annually—equivalent to about 0.1% of anthropogenic emissions—but such projections carry high uncertainty due to unverified bloom responses and competition with other nutrient sources.[94][95] Empirical validation of the whale pump's magnitude remains limited, with field observations of fecal plume effects confined to localized studies and reliant on correlations rather than causation; critics argue that enhanced primary production may not proportionally increase export flux, as much fixed carbon is respired near-surface, and global extrapolations overestimate impacts relative to the baseline biological pump's 5-12 GtC/year export. Peer-reviewed assessments emphasize that while mechanisms are plausible, rigorous quantification requires integrated modeling of whale densities, migration patterns, and nutrient stoichiometry, with ongoing debates over whether restoration efforts like whaling cessation yield measurable sequestration gains amid ocean acidification and warming stressors.[91][96]Mixoplankton and Jelly Falls
Mixoplankton, defined as planktonic organisms capable of both autotrophy through photosynthesis and heterotrophy via phagocytosis, constitute a significant portion of marine microbial communities, often comprising over 50% of protist biomass in certain oceanic regions.[97] These organisms enhance the efficiency of the biological carbon pump by bridging trophic levels, facilitating greater biomass transfer to larger grazers that produce rapidly sinking fecal pellets, thereby increasing vertical carbon flux compared to purely phototrophic or heterotrophic systems.[98] Modeling studies indicate that incorporating mixotrophy raises mean organism size and trophic transfer efficiency, potentially elevating net carbon export by up to 30% in mixotroph-dominated ecosystems.[97] For instance, in the subtropical Atlantic, mixotrophs account for 40-95% of primary production while competing with strict phytoplankton, altering carbon partitioning and export pathways through selective foraging that favors nutrient retention and particle aggregation.[99] Jelly falls refer to the mass sinking of gelatinous zooplankton carcasses, such as those from jellyfish and salps, which rapidly transport organic carbon from surface waters to the deep ocean and seafloor.[100] These events occur following blooms or die-offs, with sinking speeds exceeding 1000 meters per day for some species, far surpassing typical marine snow rates and yielding transfer efficiencies of 38-62% to 1000 meters depth.[101] Globally, gelatinous zooplankton mediate an estimated 3.7-6.8 billion metric tons of organic carbon export annually, comparable to annual anthropogenic emissions from major economies, by packaging low-density biomass into dense, ballast-enhanced aggregates that resist remineralization during descent.[102] This mechanism supplements traditional export via diatoms or fecal pellets, particularly in stratified or low-productivity waters where gelatinous blooms prevail, though their episodic nature introduces variability in pump efficiency.[100] Empirical observations from regions like the Mediterranean confirm elevated benthic carbon and nitrogen fluxes during jelly falls, underscoring their role in fueling deep-sea communities.[103]Lipid Pump from Migrations
The lipid pump refers to a biological sequestration mechanism in which vertically migrating zooplankton transport carbon-rich lipids from surface waters to mesopelagic and deeper ocean layers during seasonal overwintering diapause, contributing to long-term carbon storage with minimal nutrient recycling.[104] Unlike passive sinking of particulate organic carbon in the traditional biological pump, this process involves active migration of lipid-laden organisms to depths of 600–3,000 m in autumn, where they enter dormancy and gradually respire the stored lipids, releasing primarily carbon dioxide below the permanent thermocline.[104][105] The lipids, primarily wax esters, provide buoyancy and metabolic fuel, enabling efficient downward carbon flux without substantial energy expenditure for ascent, as the organisms remain deep until spring.[104][106] Key contributors are calanoid copepods of the genus Calanus, particularly C. finmarchicus in the North Atlantic and C. hyperboreus in Arctic and Nordic Seas regions.[104][105] These species accumulate lipids comprising over 50% of their dry weight—up to 200–300 µg per individual in late copepodite stages—during summer feeding in nutrient-replete surface layers.[104][106] Migration depths vary regionally, with C. hyperboreus reaching 1,000–3,000 m in areas like the Fram Strait and Greenland Sea, while C. finmarchicus typically overwinters at 600–1,400 m in basins such as the Labrador and Norwegian Seas.[105][106] Respiration during diapause occurs at low rates (0.4–0.8 µg C individual⁻¹ day⁻¹), sustaining the population through winter with limited remineralization until potential spring ascent or mortality.[104] Quantitatively, the lipid pump exports 2–6 g C m⁻² year⁻¹ in the North Atlantic, with 1–4 g C m⁻² year⁻¹ effectively sequestered after accounting for respiration losses, rates comparable to sinking particulate fluxes (2–8 g C m⁻² year⁻¹).[104] In the Fram Strait, C. hyperboreus alone contributes 3–6 g C m⁻² year⁻¹ at 1,000–3,000 m depths, while C. finmarchicus adds 0.3–2 Mt C year⁻¹ across the strait, with a global estimate for the species of 19.3 Mt C year⁻¹.[105][106] Inclusion of this mechanism nearly doubles prior estimates of biological carbon sequestration in the North Atlantic basin, potentially equaling 1–2.7 Gt C year⁻¹ when integrated with other pathways.[104] The efficiency of the lipid pump stems from its "lipid shunt," where stored lipids have a high carbon-to-nutrient ratio and negligible nitrogen or phosphorus content, preventing significant upward nutrient flux upon remineralization and thus avoiding surface nutrient depletion.[104][106] This decouples carbon export from nutrient cycling, contrasting with the traditional pump's reliance on nutrient-rich detritus that remineralizes shallower and replenishes surface stocks.[104] Regional variations depend on copepod abundance, lipid accumulation success, and mortality during diapause, with higher fluxes in eastern Nordic Seas due to greater biomass.[105] Potential ecological feedbacks include reduced surface primary production if lipid-stored carbon is not recycled, though empirical data remain limited to modeling and field lipid content analyses.[106]Interactions with Physical Pumps
Solubility Pump Synergies
The solubility pump and biological pump exhibit synergies that amplify overall oceanic carbon sequestration by leveraging physical transport to support biological export and vice versa. The solubility pump physically conveys dissolved inorganic carbon (DIC) from surface to deep waters through the sinking of cold, dense polar waters, where CO₂ solubility is higher due to lower temperatures; this process maintains elevated DIC concentrations in deep reservoirs, with upwelling subsequently supplying carbon and associated nutrients to surface ecosystems.[107] This influx enables enhanced phytoplankton primary production, the foundational mechanism of the biological pump, which fixes atmospheric CO₂ into organic matter at rates of approximately 10 gigatons of carbon per year globally.[107] Without the solubility pump's role in redistributing DIC, surface carbon availability for biological uptake would diminish, reducing export efficiency.[4] In turn, the biological pump augments the solubility pump by depleting surface DIC through photosynthesis, which lowers surface pCO₂ and steepens the air-sea gradient for CO₂ invasion, particularly in cold, high-solubility regions like the Southern Ocean.[7] Remineralization of sinking organic particles in the deep ocean releases DIC without equivalent alkalinity, enriching the deep DIC pool and enhancing the solubility pump's capacity to store carbon over millennial timescales; model analyses attribute about 75% of the ocean's vertical DIC gradient to biological processes, with solubility contributing 25%.[108] These feedbacks are evident in the combined contribution of both pumps to absorbing roughly 30% of anthropogenic CO₂ emissions, totaling 2.6 gigatons of carbon annually from 2004 to 2013.[107] Climate-driven changes, such as ocean warming and increased stratification, threaten these synergies by reducing solubility in surface waters and limiting nutrient upwelling, potentially weakening biological export and deep DIC replenishment.[7] Observations and models suggest that in a warmer climate, the biological pump's efficiency in high-latitude zones—where solubility is strongest—could decline, leading to feedbacks that diminish total sequestration unless offset by other dynamics.[109]Carbonate and Shelf Pump Distinctions
The carbonate pump operates through the biological production of particulate inorganic carbon (PIC) in the form of calcium carbonate (CaCO3) shells by planktonic calcifiers, such as coccolithophores and foraminifera, primarily in open ocean surface waters.[110] Calcification consumes bicarbonate ions (HCO3-), releasing CO2 proportionally, which elevates partial pressure of CO2 (pCO2) in surface waters and counteracts the atmospheric CO2 drawdown achieved by the organic biological pump.[111] Export of this PIC to depth via sinking particles contributes to long-term carbon storage upon dissolution in undersaturated deep waters, where it increases total alkalinity but does not directly sequester carbon equivalent to organic matter remineralization.[110] Global PIC export fluxes are estimated at approximately 0.4–1.6 Pg C yr-1, with coccolithophores like Emiliania huxleyi dominating production in oligotrophic gyres.[111] In contrast, the continental shelf pump describes a regionally confined mechanism on continental margins where shelf waters exhibit net CO2 uptake due to high biological productivity fueled by terrestrial nutrient inputs and stratification, leading to DIC enrichment relative to atmospheric equilibrium.[112] This excess DIC is then advected offshore to the open ocean interior via physical processes such as downwelling, eddies, and boundary currents, effectively sequestering carbon on centennial timescales beyond the shelf's rapid exchange with the atmosphere.[113] Observations in regions like the East China Sea indicate shelf sea CO2 absorption rates of about 0.02–0.03 mol m-2 day-1, with global shelf contributions potentially accounting for 0.1–0.2 Pg C yr-1 of net export to the ocean interior.[112] [113] Key distinctions between the two lie in their spatial scope, carbon form, and driving mechanisms: the carbonate pump facilitates vertical flux of PIC through biological shell formation and gravitational settling across the open ocean, influencing DIC speciation and alkalinity gradients globally, whereas the shelf pump emphasizes horizontal DIC transport from biologically productive margins, dependent on coastal circulation patterns rather than particle sinking.[110] [112] The carbonate process inherently releases CO2 during production, acting as a counterbalance to organic carbon export efficiency, while the shelf pump enhances overall oceanic CO2 uptake by isolating shelf-derived DIC from surface equilibration.[111] Both interact with the biological pump but differ in their net effect on surface pCO2: carbonate export diminishes drawdown potential, whereas shelf export amplifies sequestration via geographic separation.[113] Uncertainties persist in quantifying shelf pump strength due to variability in coastal metabolism and hydrodynamics, contrasting with more constrained PIC flux measurements from sediment traps and satellite-derived calcification rates.[113]Quantification and Efficiency
Measurement Techniques
Sediment traps, deployed as moored or neutrally buoyant devices, directly measure sinking particulate organic carbon (POC) fluxes by capturing particles at depths such as 100–150 meters below the euphotic zone, providing estimates of export from surface waters.[114] These instruments quantify the gravitational pathway of the biological pump but require corrections for hydrodynamic biases, swimmer interference from zooplankton, and dissolution effects to ensure accuracy.[14] Complementary radionuclide tracers like thorium-234 disequilibrium assess particle interception rates over timescales of weeks, often calibrating trap data by revealing flux attenuation in the upper mesopelagic zone.[115] Geochemical proxies offer indirect quantification of export and remineralization. Apparent oxygen utilization (AOU), derived from in-situ oxygen profiles on autonomous platforms like Argo floats, infers organic carbon respiration in subsurface waters via stoichiometric relationships (e.g., Redfield ratio of C:O2 ≈ 106:138), enabling basin-scale pump efficiency estimates.[116] Nutrient deficits, calculated as deviations from preformed concentrations of nitrate or phosphate, trace net export production, with subsurface maxima indicating sinking and decomposition.[10] Stable isotopes, including triple oxygen isotopes (17O/18O) and O2/Ar ratios, constrain gross primary production and net community production, distinguishing biological export from physical mixing.[117] Remote sensing techniques from satellites estimate pump components by deriving surface primary production from chlorophyll-a concentrations via algorithms like those in NASA's Ocean Biogeochemical Model, then applying empirically derived export ratios (e.g., 10–20% of net primary production) to infer carbon transfer to depth.[8] In-situ incubations with 15N-labeled nitrate measure new production as a proxy for export potential, integrating nitrate uptake rates over hours to days.[10] These methods, often combined in data-assimilative models, address spatial heterogeneity but face uncertainties from remineralization variability and regional parameterization differences.[118]Global Estimates and Historical Trends
Global estimates of the biological carbon pump's export flux, representing particulate organic carbon sinking below the euphotic zone (typically ~100 m), range from 5 to more than 12 GtC per year based on syntheses of satellite observations, sediment traps, and biogeochemical models.[119] [120] A 2024 analysis of particle flux attenuation refines the annual transfer to the ocean interior at 5–10 GtC, emphasizing the role of remineralization in limiting deep sequestration.[18] These figures derive from primary production rates of ~50 GtC yr⁻¹, with export efficiency (export-to-production ratio) averaging 10–20% globally, though higher in nutrient-rich high-latitude regions.[121] Paleoceanographic proxies, including carbon isotopes (δ¹³C) in benthic foraminifera and sediment opal fluxes, reveal enhanced biological pump efficiency during glacial maxima, such as the Last Glacial Maximum ~21,000–18,000 years ago, when atmospheric CO₂ was ~80–100 ppm lower than preindustrial levels.[122] [123] This strengthening, estimated to sequester an additional ~20–40 GtC in the ocean interior relative to interglacials, stemmed from dust-mediated iron fertilization boosting Southern Ocean productivity, reduced deep-ocean ventilation, and more complete nutrient drawdown at high latitudes.[124] [125] Over the Holocene (~11,700 years ago to present), proxy records indicate relative stability in pump strength, with fluctuations tied to orbital forcing and regional upwelling changes rather than monotonic trends.[124] In the modern era (post-1950s), direct measurements from programs like the Joint Global Ocean Flux Study show no statistically significant global decline or increase in export flux, despite rising sea surface temperatures and stratification; any anthropogenic influence remains below detection thresholds, with the solubility pump dominating CO₂ uptake.[126] [109] Model projections for the 21st century anticipate a 10–20% reduction under high-emission scenarios due to nutrient trapping in stratified surface waters, though empirical validation is limited.[127]Efficiency Metrics and Challenges
Export efficiency, defined as the fraction of net primary production exported from the euphotic zone as particulate organic carbon (POC), typically ranges from 5% to 25% globally, with higher values in nutrient-rich upwelling regions and lower in oligotrophic gyres.[114] Transfer efficiency, the proportion of POC flux at the base of the euphotic zone (around 100-150 m) that reaches the mesopelagic zone (e.g., 500-1000 m), averages 10-20% but exhibits strong regional variability, influenced by particle remineralization rates and aggregation processes.[114] Sequestration efficiency, the product of export and transfer efficiencies, quantifies the fraction of surface primary production sequestered in the deep ocean (>1000 m) for centuries, estimated at 0.1-1% globally, underscoring the pump's role in long-term carbon storage of approximately 1700 Pg C in the ocean interior.[128] These metrics are derived from field observations, including sediment trap deployments and radionuclide tracers like thorium-234, which track short-term particle export.[15] Challenges in assessing BCP efficiency stem from the heterogeneous nature of particle fluxes, where small, optically transparent particles often evade detection by traditional traps, leading to underestimates of export by up to 50% in some studies.[114] Remineralization depth profiles remain poorly constrained, with models assuming exponential decay constants (Martin curve parameters) that vary by factor of 2-3 across ocean basins, complicating global extrapolations.[129] Temporal variability, driven by seasonal blooms and episodic events like jelly falls, introduces biases in snapshot measurements, while the underrepresentation of dissolved organic matter (DOM) pathways—potentially contributing 20-50% to export—further obscures total efficiency.[15] Sparse deep-ocean observations, limited to a few long-term sites, hinder robust global budgets, with inverse modeling approaches revealing discrepancies of 20-30% between observational and simulated efficiencies.[130]- Key Metrics Summary:
Metric Definition Typical Global Range Primary Influences Export Efficiency (e-ratio) Exported POC / Net Primary Production 5-25% Nutrient availability, food web structure[114] Transfer Efficiency Flux at depth / Surface export flux 10-20% Ballast minerals, microbial degradation[129] Sequestration Efficiency Deep export (>1000 m) / Net Primary Production 0.1-1% Combined remineralization and circulation[128]