Recent from talks
Microbial loop
Knowledge base stats:
Talk channels stats:
Members stats:
Microbial loop
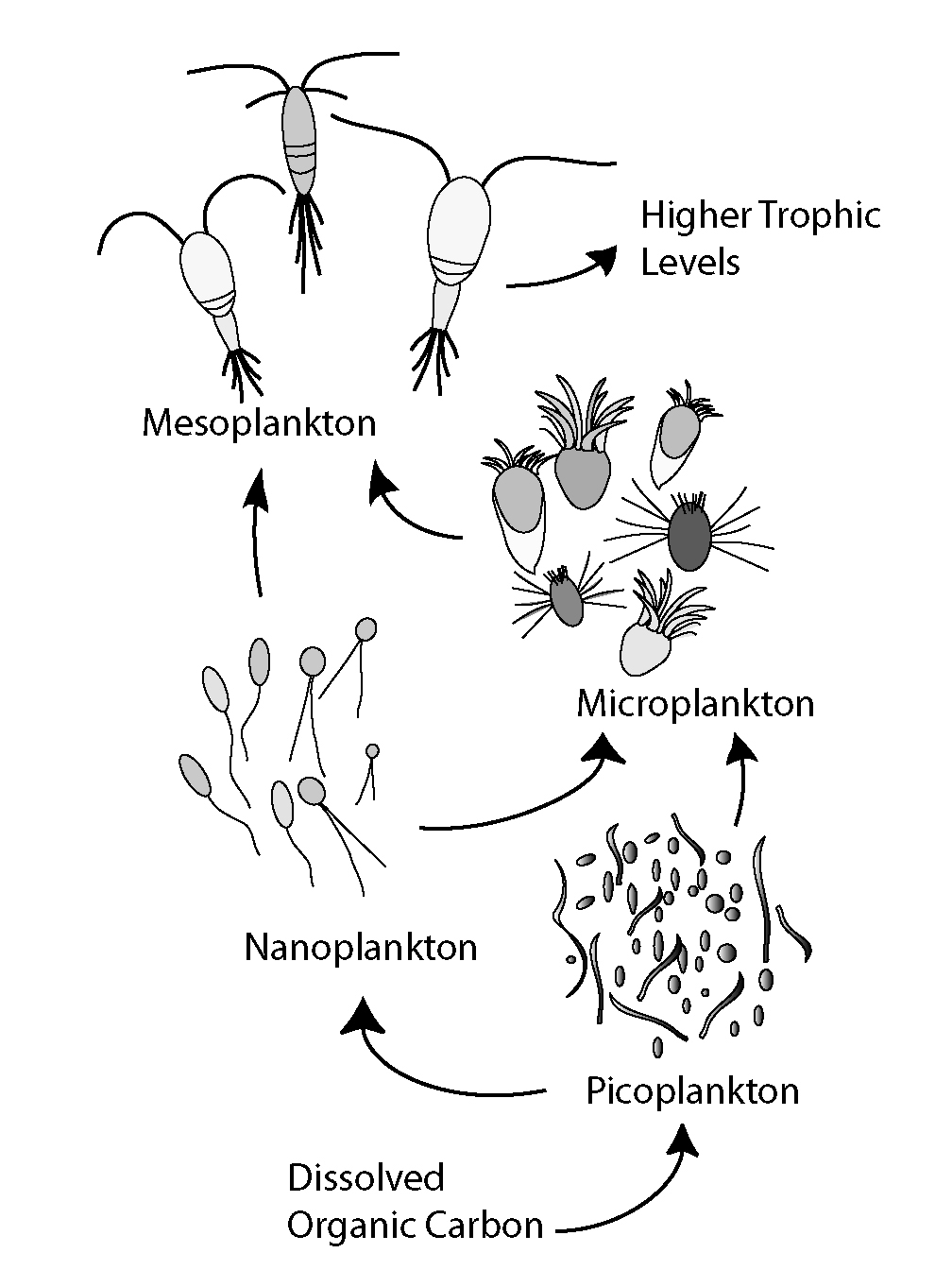
The microbial loop describes a trophic pathway where, in aquatic systems, dissolved organic carbon (DOC) is returned to higher trophic levels via its incorporation into bacterial biomass, and then coupled with the classic food chain formed by phytoplankton-zooplankton-nekton. In soil systems, the microbial loop refers to soil carbon. The term microbial loop was coined by Farooq Azam, Tom Fenchel et al. in 1983 to include the role played by bacteria in the carbon and nutrient cycles of the marine environment.
In general, dissolved organic carbon (DOC) is introduced into the ocean environment from bacterial lysis, the leakage or exudation of fixed carbon from phytoplankton (e.g., mucilaginous exopolymer from diatoms), sudden cell senescence, sloppy feeding by zooplankton, the excretion of waste products by aquatic animals, or the breakdown or dissolution of organic particles from terrestrial plants and soils. Bacteria in the microbial loop decompose this particulate detritus to utilize this energy-rich matter for growth. Since more than 95% of organic matter in marine ecosystems consists of polymeric, high-molecular- weight (HMW) compounds (e.g., proteins, polysaccharides, lipids), only a small portion of total dissolved organic matter (DOM) is readily utilizable to most marine organisms at higher trophic levels. This means that dissolved organic carbon is not available directly to most marine organisms; marine bacteria introduce this organic carbon into the food web, resulting in additional energy becoming available to higher trophic levels. Recently the term "microbial food web" has been substituted for the term "microbial loop".
Prior to the discovery of the microbial loop, the classic view of marine food webs was one of a linear chain from phytoplankton to nekton. Generally, marine bacteria were not thought to be significant consumers of organic matter (including carbon), although they were known to exist. However, the view of a marine pelagic food web was challenged during the 1970s and 1980s by Pomeroy and Azam, who suggested the alternative pathway of carbon flow from bacteria to protozoans to metazoans.
Early work in marine ecology that investigated the role of bacteria in oceanic environments concluded their role to be very minimal. Traditional methods of counting bacteria (e.g., culturing on agar plates) only yielded small numbers of bacteria that were much smaller than their true ambient abundance in seawater. Developments in technology for counting bacteria have led to an understanding of the significant importance of marine bacteria in oceanic environments.
In the 1970s, the alternative technique of direct microscopic counting was developed by Francisco et al. (1973) and Hobbie et al. (1977). Bacterial cells were counted with an epifluorescence microscope, producing what is called an "acridine orange direct count" (AODC). This led to a reassessment of the large concentration of bacteria in seawater, which was found to be more than was expected (typically on the order of 1 million per milliliter). Also, development of the "bacterial productivity assay" showed that a large fraction (i.e. 50%) of net primary production (NPP) was processed by marine bacteria.
In 1974, Larry Pomeroy published a paper in BioScience entitled "The Ocean's Food Web: A Changing Paradigm", where the key role of microbes in ocean productivity was highlighted. In the early 1980s, Azam and a panel of top ocean scientists published the synthesis of their discussion in the journal Marine Ecology Progress Series entitled "The Ecological Role of Water Column Microbes in the Sea". The term 'microbial loop' was introduced in this paper, which noted that the bacteria-consuming protists were in the same size class as phytoplankton and likely an important component of the diet of planktonic crustaceans.
Evidence accumulated since this time has indicated that some of these bacterivorous protists (such as ciliates) are actually selectively preyed upon by these copepods. In 1986, Prochlorococcus, which is found in high abundance in oligotrophic areas of the ocean, was discovered by Sallie W. Chisholm, Robert J. Olson, and other collaborators (although there had been several earlier records of very small cyanobacteria containing chlorophyll b in the ocean Prochlorococcus was discovered in 1986). Stemming from this discovery, researchers observed the changing role of marine bacteria along a nutrient gradient from eutrophic to oligotrophic areas in the ocean.
The efficiency of the microbial loop is determined by the density of marine bacteria within it. It has become clear that bacterial density is mainly controlled by the grazing activity of small protozoans such as various taxonomic groups of flagellates. Also, viral infection causes bacterial lysis, which release cell contents back into the dissolved organic matter (DOM) pool, lowering the overall efficiency of the microbial loop. Mortality from viral infection has almost the same magnitude as that from protozoan grazing. However, compared to protozoan grazing, the effect of viral lysis can be very different because lysis is highly host-specific to each marine bacteria. Both protozoan grazing and viral infection balance the major fraction of bacterial growth. In addition, the microbial loop dominates in oligotrophic waters, rather than in eutrophic areas - there the classical plankton food chain predominates, due to the frequent fresh supply of mineral nutrients (e.g. spring bloom in temperate waters, upwelling areas). The magnitude of the efficiency of the microbial loop can be determined by measuring bacterial incorporation of radiolabeled substrates (such as tritiated thymidine or leucine).
Hub AI
Microbial loop AI simulator
(@Microbial loop_simulator)
Microbial loop
The microbial loop describes a trophic pathway where, in aquatic systems, dissolved organic carbon (DOC) is returned to higher trophic levels via its incorporation into bacterial biomass, and then coupled with the classic food chain formed by phytoplankton-zooplankton-nekton. In soil systems, the microbial loop refers to soil carbon. The term microbial loop was coined by Farooq Azam, Tom Fenchel et al. in 1983 to include the role played by bacteria in the carbon and nutrient cycles of the marine environment.
In general, dissolved organic carbon (DOC) is introduced into the ocean environment from bacterial lysis, the leakage or exudation of fixed carbon from phytoplankton (e.g., mucilaginous exopolymer from diatoms), sudden cell senescence, sloppy feeding by zooplankton, the excretion of waste products by aquatic animals, or the breakdown or dissolution of organic particles from terrestrial plants and soils. Bacteria in the microbial loop decompose this particulate detritus to utilize this energy-rich matter for growth. Since more than 95% of organic matter in marine ecosystems consists of polymeric, high-molecular- weight (HMW) compounds (e.g., proteins, polysaccharides, lipids), only a small portion of total dissolved organic matter (DOM) is readily utilizable to most marine organisms at higher trophic levels. This means that dissolved organic carbon is not available directly to most marine organisms; marine bacteria introduce this organic carbon into the food web, resulting in additional energy becoming available to higher trophic levels. Recently the term "microbial food web" has been substituted for the term "microbial loop".
Prior to the discovery of the microbial loop, the classic view of marine food webs was one of a linear chain from phytoplankton to nekton. Generally, marine bacteria were not thought to be significant consumers of organic matter (including carbon), although they were known to exist. However, the view of a marine pelagic food web was challenged during the 1970s and 1980s by Pomeroy and Azam, who suggested the alternative pathway of carbon flow from bacteria to protozoans to metazoans.
Early work in marine ecology that investigated the role of bacteria in oceanic environments concluded their role to be very minimal. Traditional methods of counting bacteria (e.g., culturing on agar plates) only yielded small numbers of bacteria that were much smaller than their true ambient abundance in seawater. Developments in technology for counting bacteria have led to an understanding of the significant importance of marine bacteria in oceanic environments.
In the 1970s, the alternative technique of direct microscopic counting was developed by Francisco et al. (1973) and Hobbie et al. (1977). Bacterial cells were counted with an epifluorescence microscope, producing what is called an "acridine orange direct count" (AODC). This led to a reassessment of the large concentration of bacteria in seawater, which was found to be more than was expected (typically on the order of 1 million per milliliter). Also, development of the "bacterial productivity assay" showed that a large fraction (i.e. 50%) of net primary production (NPP) was processed by marine bacteria.
In 1974, Larry Pomeroy published a paper in BioScience entitled "The Ocean's Food Web: A Changing Paradigm", where the key role of microbes in ocean productivity was highlighted. In the early 1980s, Azam and a panel of top ocean scientists published the synthesis of their discussion in the journal Marine Ecology Progress Series entitled "The Ecological Role of Water Column Microbes in the Sea". The term 'microbial loop' was introduced in this paper, which noted that the bacteria-consuming protists were in the same size class as phytoplankton and likely an important component of the diet of planktonic crustaceans.
Evidence accumulated since this time has indicated that some of these bacterivorous protists (such as ciliates) are actually selectively preyed upon by these copepods. In 1986, Prochlorococcus, which is found in high abundance in oligotrophic areas of the ocean, was discovered by Sallie W. Chisholm, Robert J. Olson, and other collaborators (although there had been several earlier records of very small cyanobacteria containing chlorophyll b in the ocean Prochlorococcus was discovered in 1986). Stemming from this discovery, researchers observed the changing role of marine bacteria along a nutrient gradient from eutrophic to oligotrophic areas in the ocean.
The efficiency of the microbial loop is determined by the density of marine bacteria within it. It has become clear that bacterial density is mainly controlled by the grazing activity of small protozoans such as various taxonomic groups of flagellates. Also, viral infection causes bacterial lysis, which release cell contents back into the dissolved organic matter (DOM) pool, lowering the overall efficiency of the microbial loop. Mortality from viral infection has almost the same magnitude as that from protozoan grazing. However, compared to protozoan grazing, the effect of viral lysis can be very different because lysis is highly host-specific to each marine bacteria. Both protozoan grazing and viral infection balance the major fraction of bacterial growth. In addition, the microbial loop dominates in oligotrophic waters, rather than in eutrophic areas - there the classical plankton food chain predominates, due to the frequent fresh supply of mineral nutrients (e.g. spring bloom in temperate waters, upwelling areas). The magnitude of the efficiency of the microbial loop can be determined by measuring bacterial incorporation of radiolabeled substrates (such as tritiated thymidine or leucine).
Recent media